

Mitochondrial ROS signaling in organismal homeostasis
- PMID: 26496603
- PMCID: PMC4634671
- DOI: 10.1016/j.cell.2015.10.001
Mitochondrial ROS signaling in organismal homeostasis
Abstract
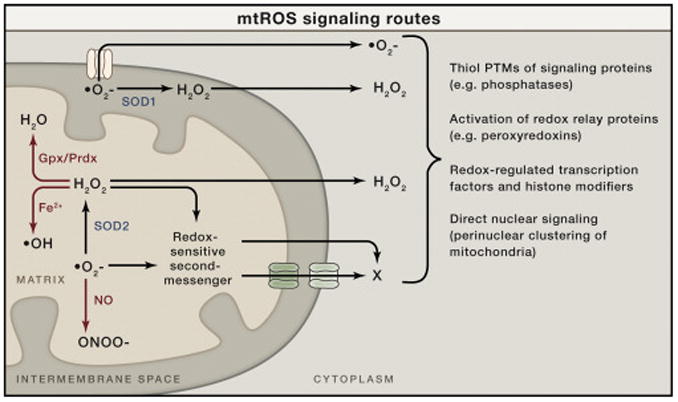
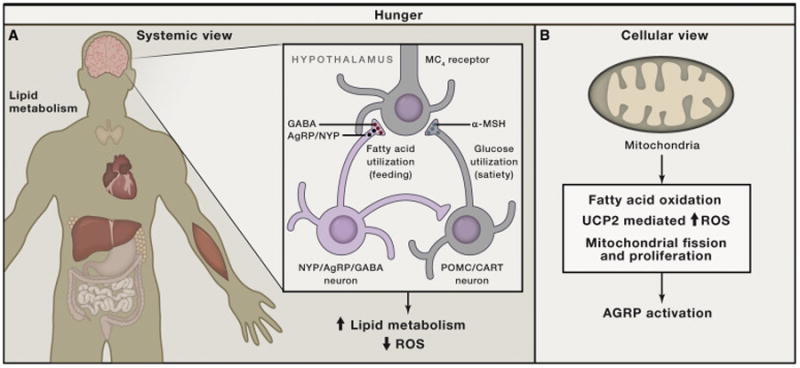
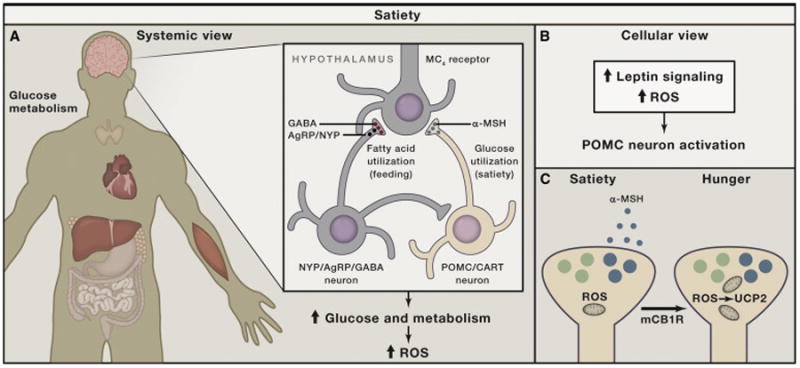
Generation, transformation, and utilization of organic molecules in support of cellular differentiation, growth, and maintenance are basic tenets that define life. In eukaryotes, mitochondrial oxygen consumption plays a central role in these processes. During the process of oxidative phosphorylation, mitochondria utilize oxygen to generate ATP from organic fuel molecules but in the process also produce reactive oxygen species (ROS). While ROS have long been appreciated for their damage-promoting, detrimental effects, there is now a greater understanding of their roles as signaling molecules. Here, we review mitochondrial ROS-mediated signaling pathways with an emphasis on how they are involved in various basal and adaptive physiological responses that control organismal homeostasis.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
