

Backtracking behavior in viral RNA-dependent RNA polymerase provides the basis for a second initiation site
- PMID: 26496948
- PMCID: PMC4666362
- DOI: 10.1093/nar/gkv1098
Backtracking behavior in viral RNA-dependent RNA polymerase provides the basis for a second initiation site
Abstract
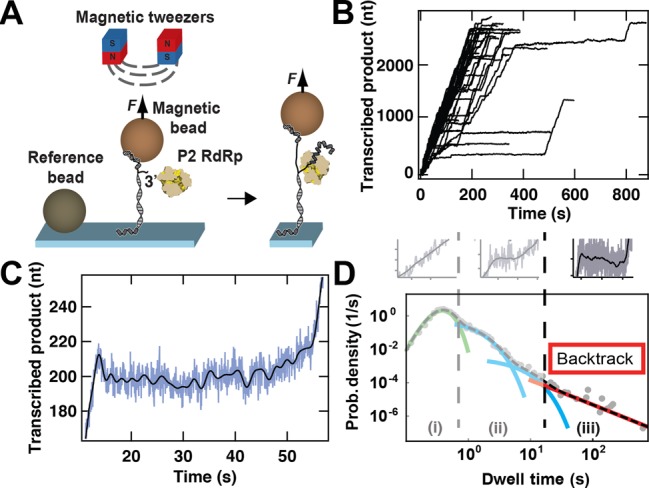
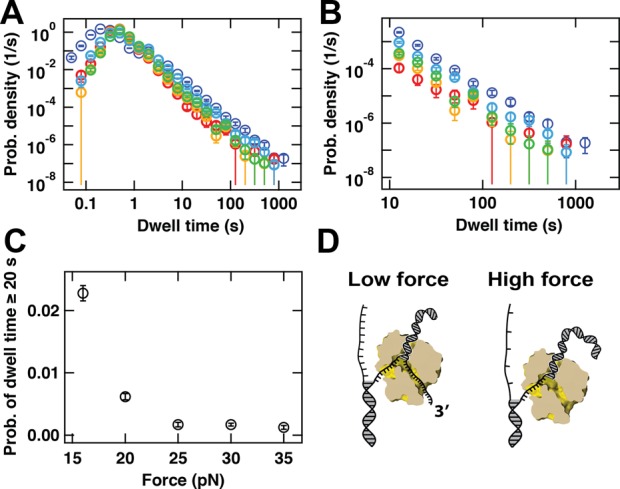
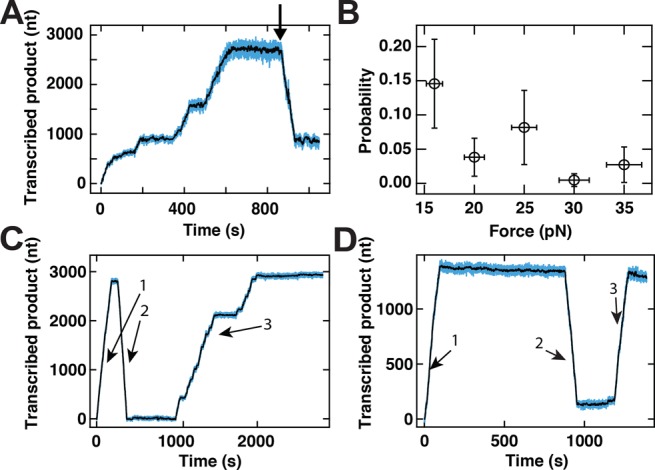
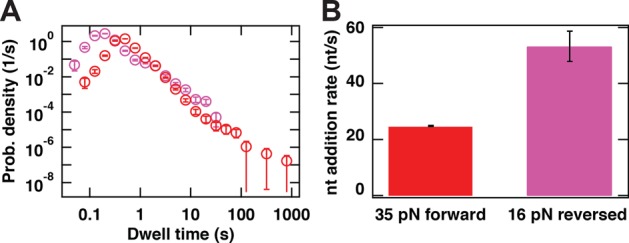
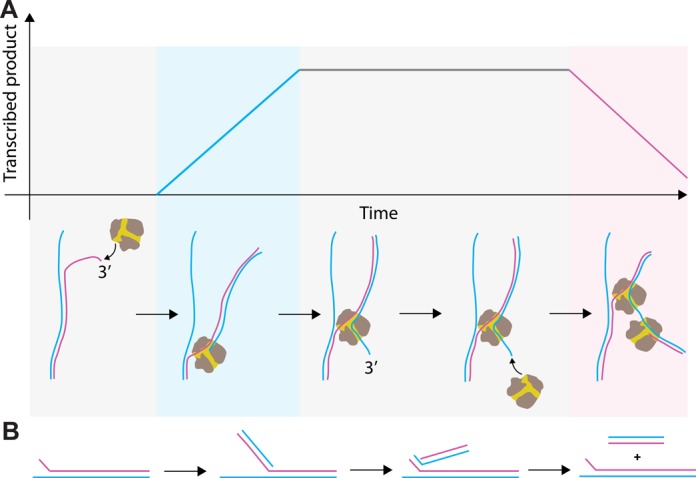
Transcription in RNA viruses is highly dynamic, with a variety of pauses interrupting nucleotide addition by RNA-dependent RNA polymerase (RdRp). For example, rare but lengthy pauses (>20 s) have been linked to backtracking for viral single-subunit RdRps. However, while such backtracking has been well characterized for multi-subunit RNA polymerases (RNAPs) from bacteria and yeast, little is known about the details of viral RdRp backtracking and its biological roles. Using high-throughput magnetic tweezers, we quantify the backtracking by RdRp from the double-stranded (ds) RNA bacteriophage Φ6, a model system for RdRps. We characterize the probability of entering long backtracks as a function of force and propose a model in which the bias toward backtracking is determined by the base paring at the dsRNA fork. We further discover that extensive backtracking provides access to a new 3'-end that allows for the de novo initiation of a second RdRp. This previously unidentified behavior provides a new mechanism for rapid RNA synthesis using coupled RdRps and hints at a possible regulatory pathway for gene expression during viral RNA transcription.
© The Author(s) 2015. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Dual Role of a Viral Polymerase in Viral Genome Replication and Particle Self-Assembly.mBio. 2018 Oct 2;9(5):e01242-18. doi: 10.1128/mBio.01242-18. mBio. 2018. PMID: 30279282 Free PMC article.
-
RNA-dependent RNA polymerases of dsRNA bacteriophages.Virus Res. 2004 Apr;101(1):45-55. doi: 10.1016/j.virusres.2003.12.005. Virus Res. 2004. PMID: 15010216 Review.
-
Temperature requirements for initiation of RNA-dependent RNA polymerization.Virology. 2003 Sep 30;314(2):706-15. doi: 10.1016/s0042-6822(03)00460-4. Virology. 2003. PMID: 14554097
-
Primer-independent RNA sequencing with bacteriophage phi6 RNA polymerase and chain terminators.RNA. 2001 May;7(5):774-81. doi: 10.1017/s1355838201002060. RNA. 2001. PMID: 11350041 Free PMC article.
-
RNA-dependent RNA polymerases in RNA silencing.Biol Chem. 2011 Apr;392(4):299-304. doi: 10.1515/BC.2011.035. Epub 2011 Feb 7. Biol Chem. 2011. PMID: 21294682 Review.
Cited by
-
Correction-free force calibration for magnetic tweezers experiments.Sci Rep. 2018 Oct 29;8(1):15920. doi: 10.1038/s41598-018-34360-4. Sci Rep. 2018. PMID: 30374099 Free PMC article.
-
RNA polymerization actuating nucleic acid membrane (RANAM)-based biosensing for universal RNA virus detection.Biosens Bioelectron. 2022 Mar 1;199:113880. doi: 10.1016/j.bios.2021.113880. Epub 2021 Dec 10. Biosens Bioelectron. 2022. PMID: 34915215 Free PMC article.
-
Force spectroscopy with electromagnetic tweezers.J Appl Phys. 2021 Oct 7;130(13):134702. doi: 10.1063/5.0060276. Epub 2021 Oct 5. J Appl Phys. 2021. PMID: 38681504 Free PMC article.
-
Characterizing single-molecule dynamics of viral RNA-dependent RNA polymerases with multiplexed magnetic tweezers.STAR Protoc. 2022 Aug 5;3(3):101606. doi: 10.1016/j.xpro.2022.101606. eCollection 2022 Sep 16. STAR Protoc. 2022. PMID: 35959497 Free PMC article.
-
Structural basis for backtracking by the SARS-CoV-2 replication-transcription complex.bioRxiv [Preprint]. 2021 Mar 14:2021.03.13.435256. doi: 10.1101/2021.03.13.435256. bioRxiv. 2021. Update in: Cell. 2020 Sep 17;182(6):1560-1573.e13. doi: 10.1016/j.cell.2020.07.033. Update in: Proc Natl Acad Sci U S A. 2021 May 11;118(19):e2102516118. doi: 10.1073/pnas.2102516118. PMID: 33758867 Free PMC article. Updated. Preprint.
References
-
- Alberts B. Molecular biology of the cell. 4th edn. NY: Garland Science; 2002.
-
- Dulin D., Lipfert J., Moolman M.C., Dekker N.H. Studying genomic processes at the single-molecule level: introducing the tools and applications. Nat. Rev. Gen. 2013;14:9–22. - PubMed
-
- Geertsema H.J., van Oijen A.M. A single-molecule view of DNA replication: the dynamic nature of multi-protein complexes revealed. Curr. Opin. Struct. Biol. 2013;23:788–793. - PubMed
-
- Lee J.B., Hite R.K., Hamdan S.M., Xie X.S., Richardson C.C., van Oijen A.M. DNA primase acts as a molecular brake in DNA replication. Nature. 2006;439:621–624. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
