

Multi-level engineering facilitates the production of phenylpropanoid compounds in tomato
- PMID: 26497596
- PMCID: PMC4639801
- DOI: 10.1038/ncomms9635
Multi-level engineering facilitates the production of phenylpropanoid compounds in tomato
Abstract
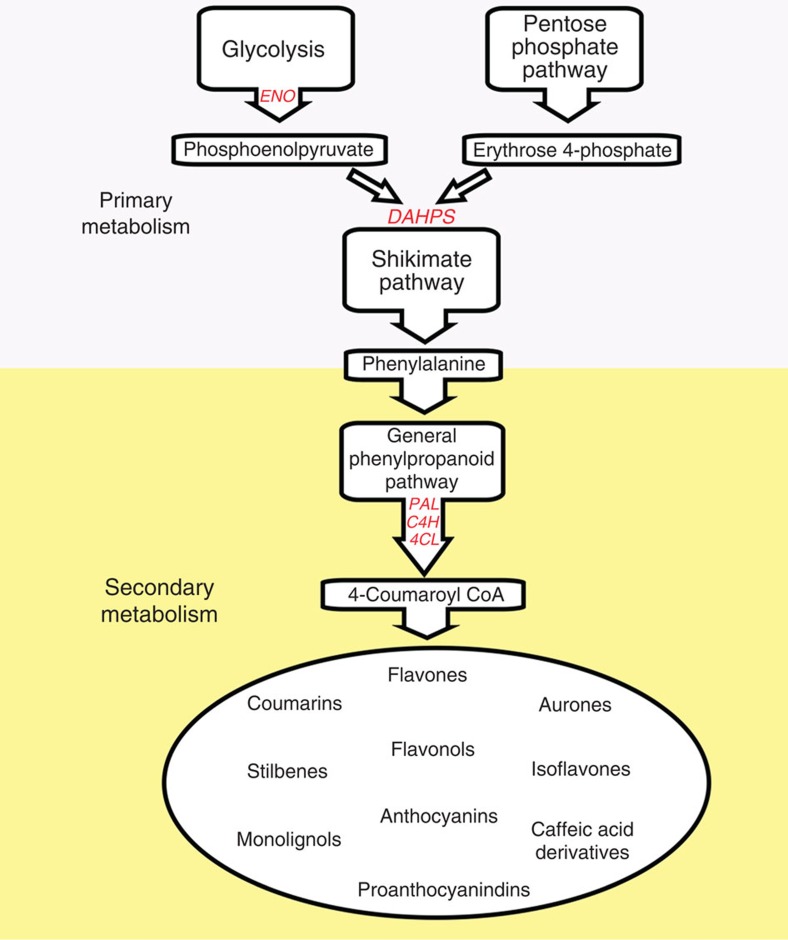
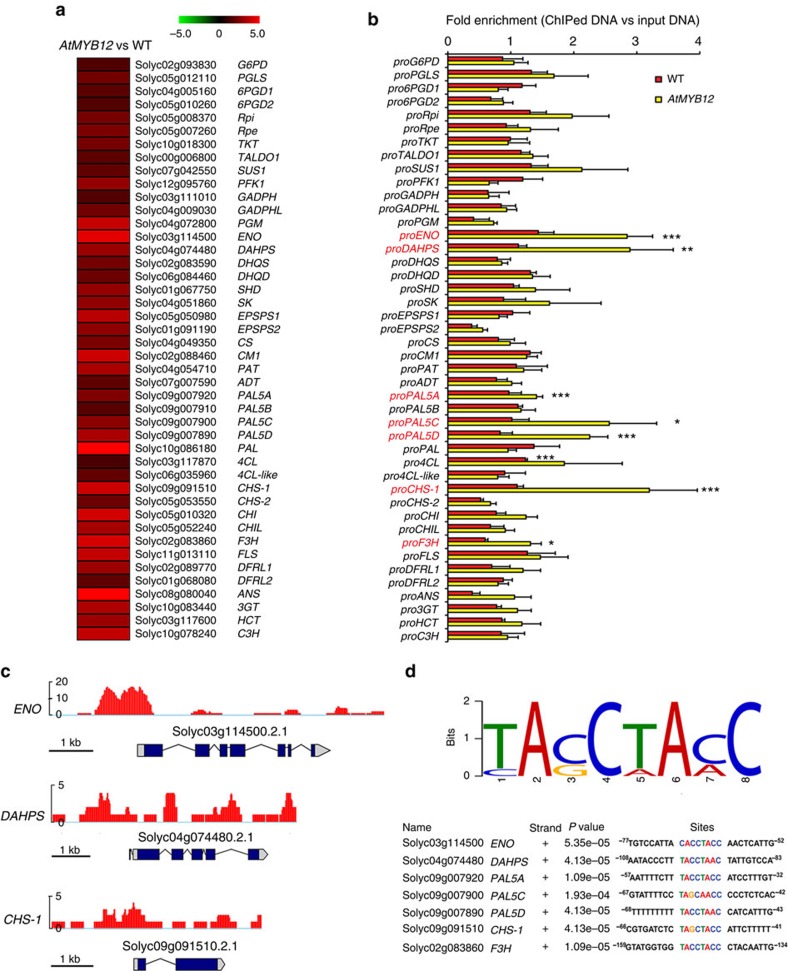
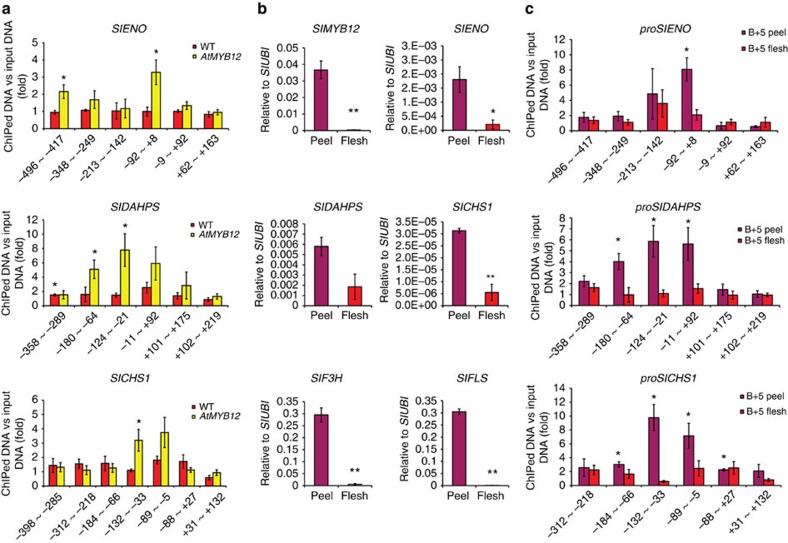
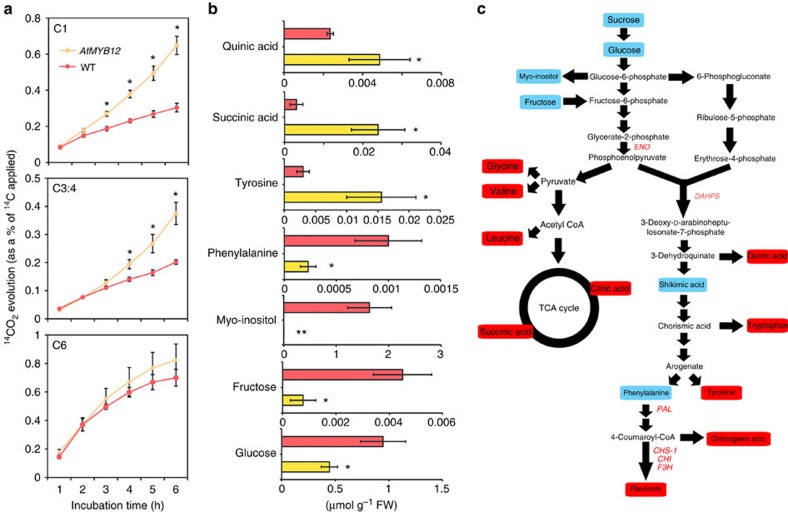
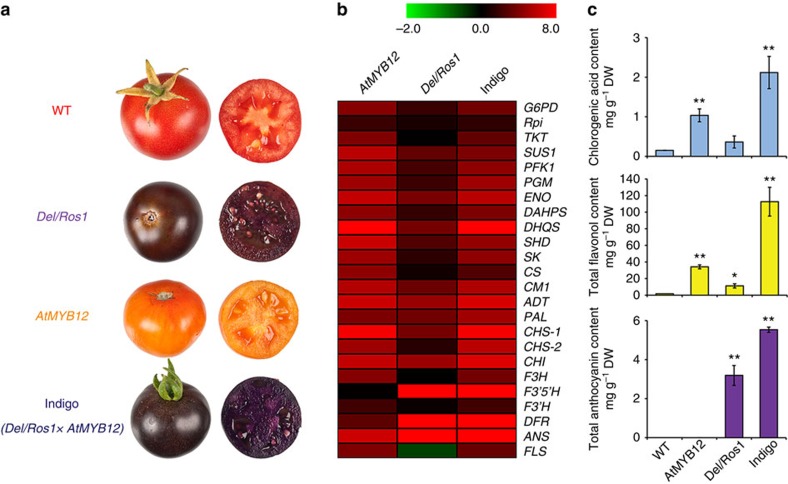
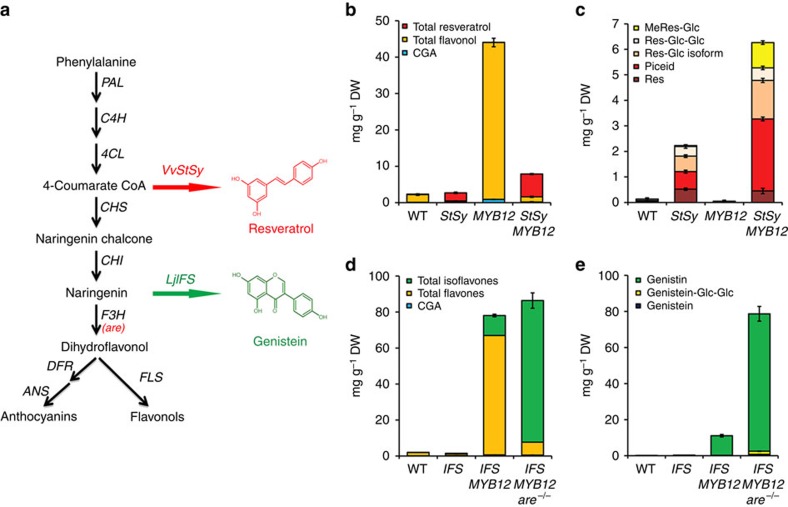
Phenylpropanoids comprise an important class of plant secondary metabolites. A number of transcription factors have been used to upregulate-specific branches of phenylpropanoid metabolism, but by far the most effective has been the fruit-specific expression of AtMYB12 in tomato, which resulted in as much as 10% of fruit dry weight accumulating as flavonols and hydroxycinnamates. We show that AtMYB12 not only increases the demand of flavonoid biosynthesis but also increases the supply of carbon from primary metabolism, energy and reducing power, which may fuel the shikimate and phenylalanine biosynthetic pathways to supply more aromatic amino acids for secondary metabolism. AtMYB12 directly binds promoters of genes encoding enzymes of primary metabolism. The enhanced supply of precursors, energy and reducing power achieved by AtMYB12 expression can be harnessed to engineer high levels of novel phenylpropanoids in tomato fruit, offering an effective production system for bioactives and other high value ingredients.
Conflict of interest statement
C.M. is an unpaid director of Norfolk Plant Sciences Ltd. E.B. and C.M. are unpaid directors of Persephone Bio Ltd. The remaining authors declare no competing financial interests.
Figures
References
-
- Butelli E. et al. Enrichment of tomato fruit with health-promoting anthocyanins by expression of select transcription factors. Nat. Biotechnol. 26, 1301–1308 (2008). - PubMed
-
- Luo J. et al. AtMYB12 regulates caffeoyl quinic acid and flavonol synthesis in tomato: expression in fruit results in very high levels of both types of polyphenol. Plant J. 56, 316–326 (2008). - PubMed
-
- Herrmann K. M. & Weaver L. M. The Shikimate Pathway. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 50, 473–503 (1999). - PubMed
-
- Tzin V. et al. Expression of a bacterial feedback-insensitive 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase of the shikimate pathway in Arabidopsis elucidates potential metabolic bottlenecks between primary and secondary metabolism. New Phytol. 194, 430–439 (2012). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
