

Arterivirus RNA-dependent RNA polymerase: Vital enzymatic activity remains elusive
- PMID: 26499043
- PMCID: PMC7111705
- DOI: 10.1016/j.virol.2015.10.002
Arterivirus RNA-dependent RNA polymerase: Vital enzymatic activity remains elusive
Abstract
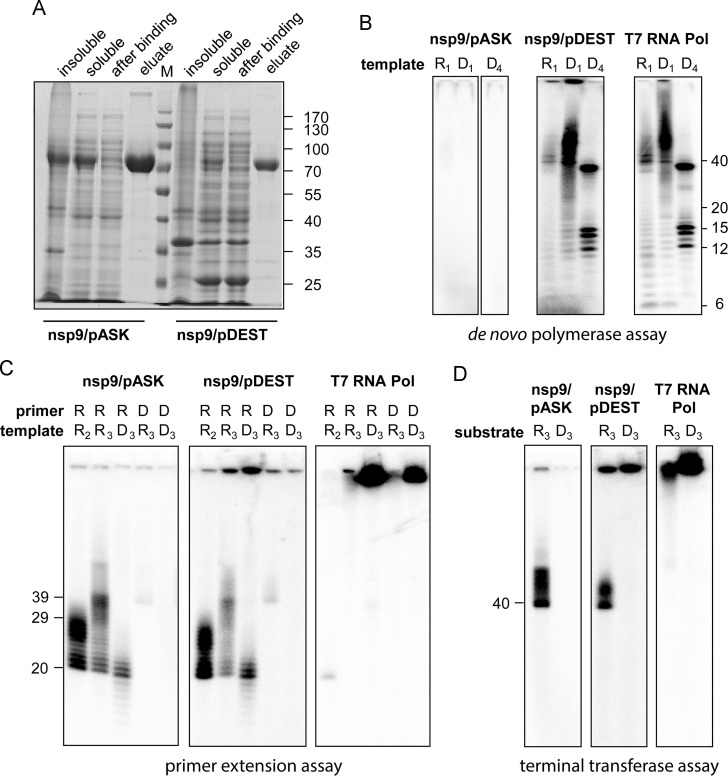
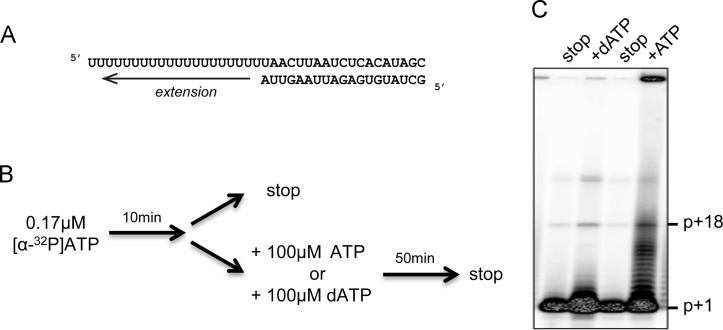
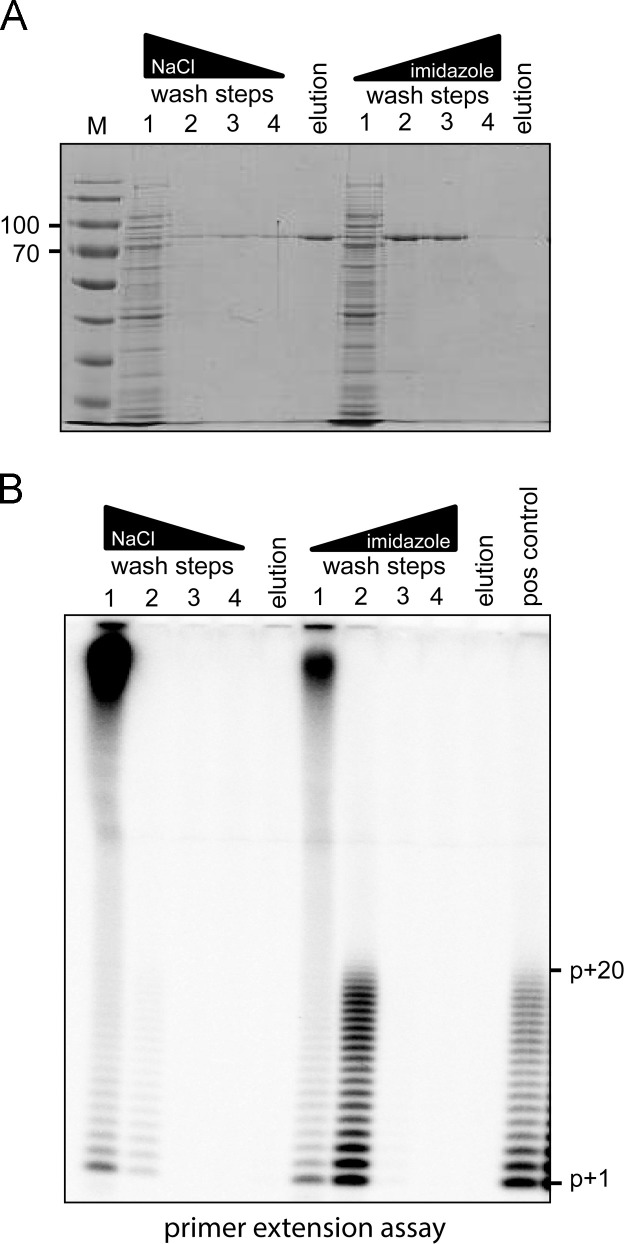
All RNA viruses encode an RNA-dependent RNA polymerase (RdRp), which in arteriviruses is expressed as the C-terminal domain of nonstructural protein 9 (nsp9). Previously, potent primer-dependent RdRp activity has been demonstrated for the homologous polymerase subunit (nsp12) of the distantly related coronaviruses. The only previous study focusing on the in vitro activity of nsp9 of an arterivirus (equine arteritis virus; EAV) reported weak de novo polymerase activity on homopolymeric RNA templates. However, this activity was not retained when Mn(2+) ions were omitted from the assay or when biologically relevant templates were supplied, which prompted us to revisit the biochemical properties of this polymerase. Based on the properties of active-site mutants, we conclude that the RNA-synthesizing activities observed in de novo and primer-dependent polymerase and terminal transferase assays cannot be attributed to recombinant EAV nsp9-RdRp. Our results illustrate the potential pitfalls of characterizing polymerases using highly sensitive biochemical assays.
Keywords: Equine arteritis virus; In vitro RdRp activity; Nidovirus; Polymerase active site mutant; Recombinant nsp9; Reverse genetics.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures

References
-
- Arnold J.J., Ghosh S.K., Cameron C.E. Poliovirus RNA-dependent RNA polymerase (3D(pol)). Divalent cation modulation of primer, template, and nucleotide selection. J. Biol. Chem. 1999;274:37060–37069. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
