

Review
doi: 10.1038/nrmicro3552.
Epub 2015 Oct 26.
Gut biogeography of the bacterial microbiota
Affiliations
- PMID: 26499895
- PMCID: PMC4837114
- DOI: 10.1038/nrmicro3552
Item in Clipboard
Review
Gut biogeography of the bacterial microbiota
Nat Rev Microbiol.
2016 Jan.
Abstract
Animals assemble and maintain a diverse but host-specific gut microbial community. In addition to characteristic microbial compositions along the longitudinal axis of the intestines, discrete bacterial communities form in microhabitats, such as the gut lumen, colonic mucus layers and colonic crypts. In this Review, we examine how the spatial distribution of symbiotic bacteria among physical niches in the gut affects the development and maintenance of a resilient microbial ecosystem. We consider novel hypotheses for how nutrient selection, immune activation and other mechanisms control the biogeography of bacteria in the gut, and we discuss the relevance of this spatial heterogeneity to health and disease.
Figures
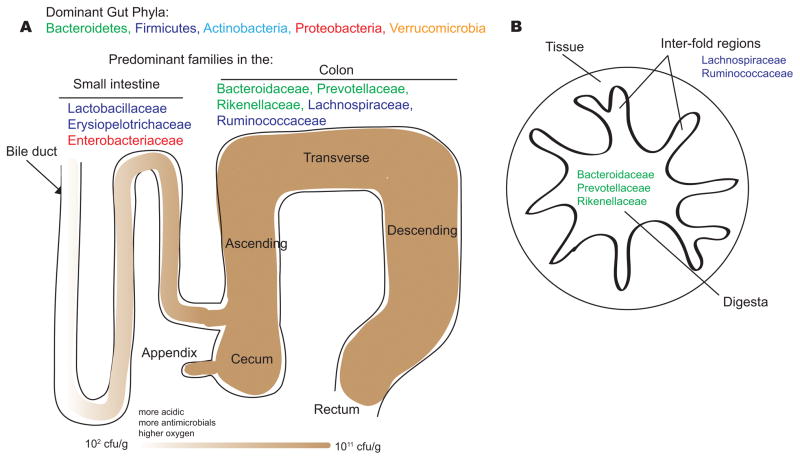
The dominant bacterial phyla in the gut are the Bacteroidetes, Firmicutes, Actinobacteria, Proteobacteria and Verrucomicrobia. The dominant bacterial families of the small intestine and colon reflect physiological differences along the length of the gut. For example, a gradient of oxygen, antimicrobial peptides (including bile acids, secreted by the bile duct), and pH limits the bacterial density in the small intestinal community, whereas the colon carries high bacterial loads. In the small intestine, Lactobacillaceae and Enterobacteriaceae dominate, whereas the colon is characterized by the presence of Bacteroidaceae, Prevotellaceae, Rikenellaceae, Lachnospiraceae and Ruminococcaceae. A cross-section of the colon shows the digesta – which is dominated by Bacteroidaceae, Prevotellaceae and Rikenellaceae – and the inter-fold regions of the lumen – which are dominated by Lachnospiraceae and Ruminococcaceae.
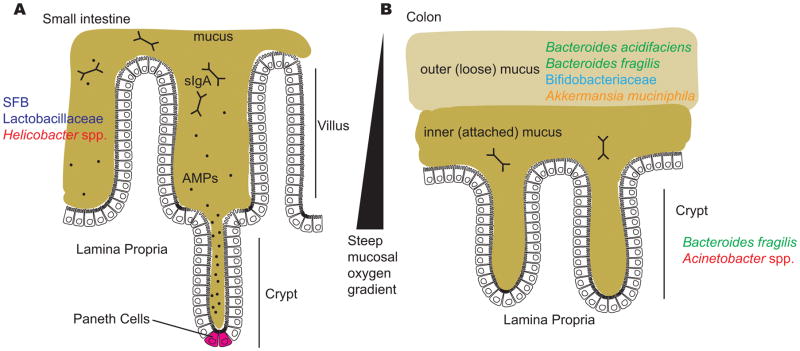
Several factors limit the ability of gut bacteria to access host cells, including the mucus layers in the small intestine and the colon; antimicrobial peptides in the small intestine, including those produced by Paneth cells at the base of the crypts; secreted immunoglobulin A (sIgA) in both the small intestine and colon; and a steep oxygen gradient that influences which bacteria are capable of surviving close to the epithelial surface. A) The surface of the small intestine is shaped into villi and crypts and is colonized by certain adherent species, including segmented filamentous bacteria (SFB), Lactobacillaceae and Helicobacter spp. B) The colon has two distinct mucus structures: the outer layer is colonized by mucin-degrading bacteria and is characterized by the presence of Bacteroides acidifaciens, Bacteroides fragilis, Bifidobacteriaceae and Akkermansia muciniphila and the inner layer and crypts are penetrated at low density by a more restricted community that includes Bacteroides fragilis and Acinetobacter spp.
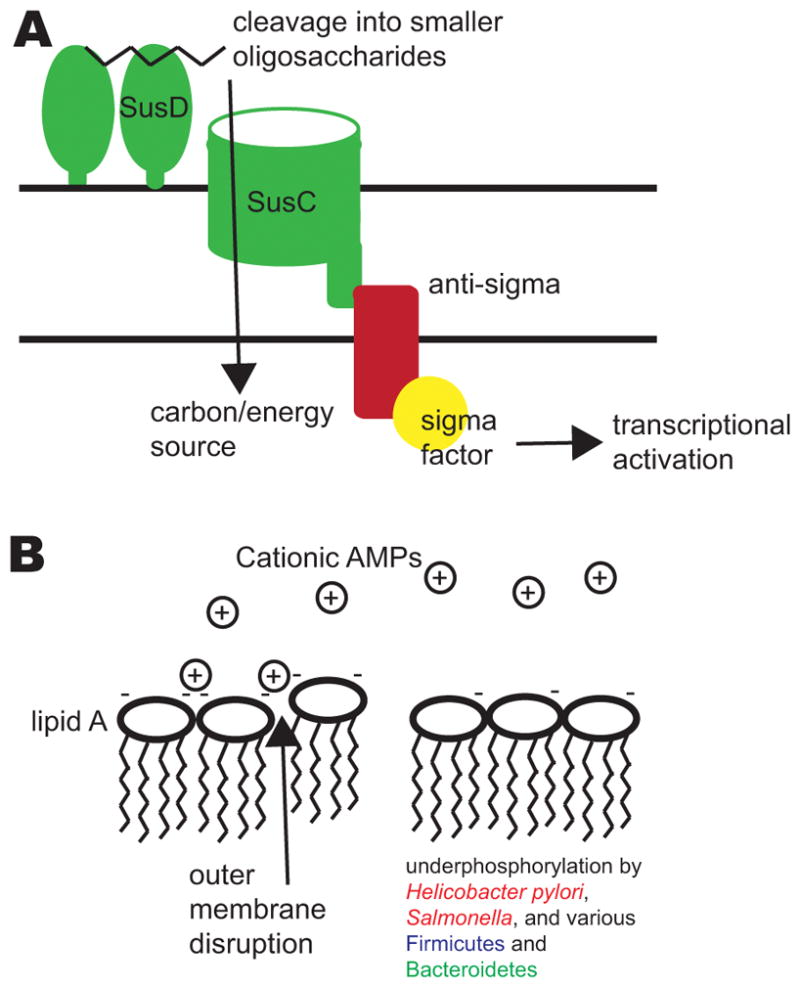
Several factors affect the localization of bacteria within the gastrointestinal tract, including the ability to utilize different glycans and to resist antimicrobial peptides (AMPs). A) Sus-like systems in Bacteroides species allow the utilization of complex polysaccharides from the diet or the host. The figure illustrates a generalized schematic of a Sus-like system. Homologues of SusD and other outer membrane lipid-anchored enzymes bind and cleave the glycans (such as starch) into smaller oligosaccharides that are then imported by the SusC-like outer membrane transporter. Interaction with the cognate glycan often leads to transmembrane signaling to activate gene regulatory mechanisms, such as a two-component system or a transmembrane anti-sigma factor which releases and activates a sigma factor. Downstream transcriptional regulation allows Bacteroides species to respond to local availability of glycans. B) Cationic AMPs in the small intestine, which also pass into the colon via the fecal stream, disrupt bacterial outer membranes by interacting with negative charges on their surface. By removing phosphate groups from lipid A of lipopolysaccharide (LPS), pathogens and commensals alike – such as Helicobacter pylori, Salmonella spp., and various Firmicutes and Bacteroidetes –reduce the negative charge on their membranes and evade attack by cationic AMPs.
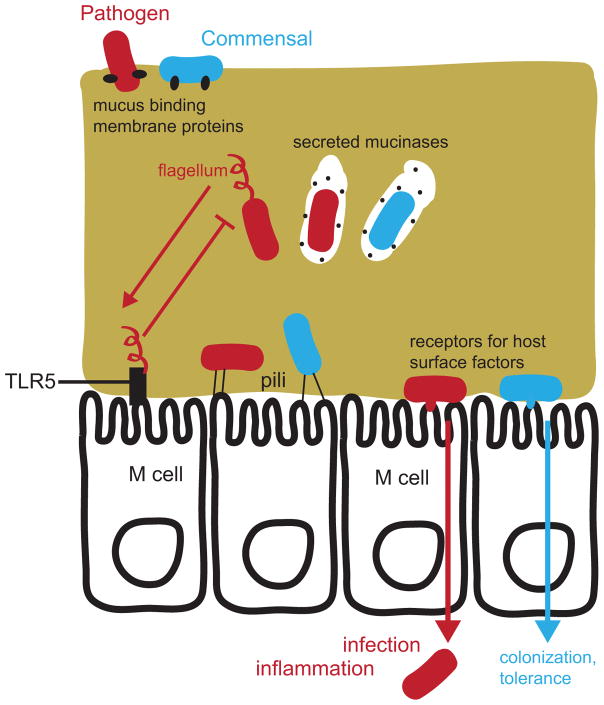
Both bacterial pathogens (red) and commensals (or mutualists; blue) have the ability to cross the mucus layer and access the gut epithelium. Lectins and other mucus-binding proteins facilitate initial interactions with the mucus layer. Mucinases and proteases are used to degrade mucus for bacteria to “eat” their way through, while some pathogens such as Salmonella spp. use flagella to swim through the viscous mucus. TLR5 sensing of flagellin effectively leads to inhibition of flagellar biosynthesis for most bacteria in the gut. Adherence to the tissue is achieved by both commensals and pathogens through pili, lectins, and other outer-membrane proteins that target ligands on the epithelial cell surface. Adherence facilitates gut colonization for both commensals and pathogens, and also allows tissue invasion by pathogenic bacteria. Microfold cells (M cells) are specialized immune sentinel epithelial cells that detect gut bacteria and are also exploited by many pathogens as a means of translocation across the epithelium.
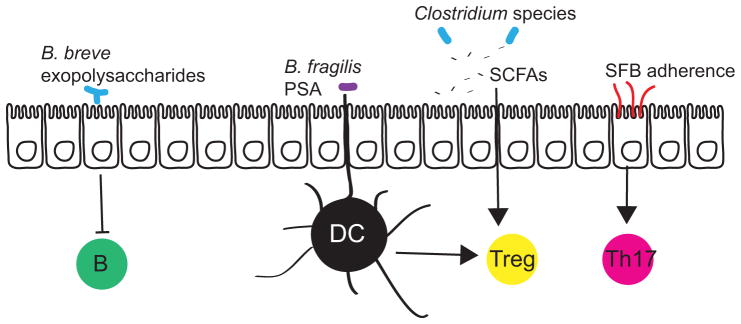
Commensal gut bacteria induce immunomodulation via interaction with epithelial cells, antigen presenting cells (such as dendritic cells (DCs)), and via production of signaling metabolites. The exopolysaccharides of adherent Bifidobacterium breve reduce the production of inflammatory cytokines to dampen B cell responses. The capsular polysaccharide PSA of Bacteroides fragilis and short-chain fatty acids (SCFAs) produced by many species of Clostridia (and other genera) stimulate the production of the anti-inflammatory interleukin-10 (IL-10) by regulatory T cells. Segmented filamentous bacteria (SFB) intercalate between microvilli of epithelial cells and stimulate the development of Th17 cells, which are important for mucosal immunity to extracellular pathogens.
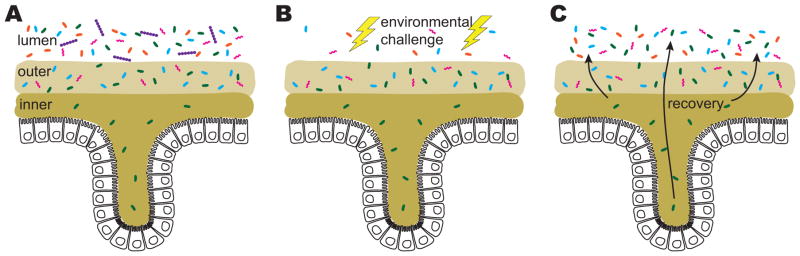
Specific niches such as crypts, the inner mucus, and the appendix may be crucial to facilitate immune homeostasis, to protect microbial inhabitants from competitors, and to re-populate the gut following perturbations that alter bacterial community structure or deplete certain species from the lumen. A) A subset of species is able to penetrate the inner mucus layer and enter crypt spaces. B) Environmental challenges such as diet perturbations, abnormalities in gastrointestinal motility, and antibiotic consumption massively alter the lumen community. However, the more stable mucosal environment and crypts protect important bacterial species. C) The crypts and mucosa serve as reservoirs to repopulate the lumen.
References
-
- Bevins CL, Salzman NH. Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Nat Rev Microbiol. 2011;9:356–368. - PubMed
-
- Peterson DA, McNulty NP, Guruge JL, Gordon JI. IgA response to symbiotic bacteria as a mediator of gut homeostasis. Cell Host Microbe. 2007;2:328–339. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
