An official website of the United States government
The .gov means it’s official.
Federal government websites often end in .gov or .mil. Before
sharing sensitive information, make sure you’re on a federal
government site.
The site is secure.
The https:// ensures that you are connecting to the
official website and that any information you provide is encrypted
and transmitted securely.
The morphogenic factor sonic hedgehog (Shh) actively orchestrates many aspects of cerebellar development and maturation. During embryogenesis, Shh signaling is active in the ventricular germinal zone (VZ) and represents an essential signal for proliferation of VZ-derived progenitors. Later, Shh secreted by Purkinje cells sustains the amplification of postnatal neurogenic niches: the external granular layer and the prospective white matter, where excitatory granule cells and inhibitory interneurons are produced, respectively. Moreover, Shh signaling affects Bergmann glial differentiation and promotes cerebellar foliation during development. Here we review the most relevant functions of Shh during cerebellar ontogenesis, underlying its role in physiological and pathological conditions.
The author(s) declare that they have no competing interests.
Figures
Fig. 1
Shh signaling pathway in vertebrates. …
Fig. 1
Shh signaling pathway in vertebrates. A 1 Shh is translated as a…
Fig. 1
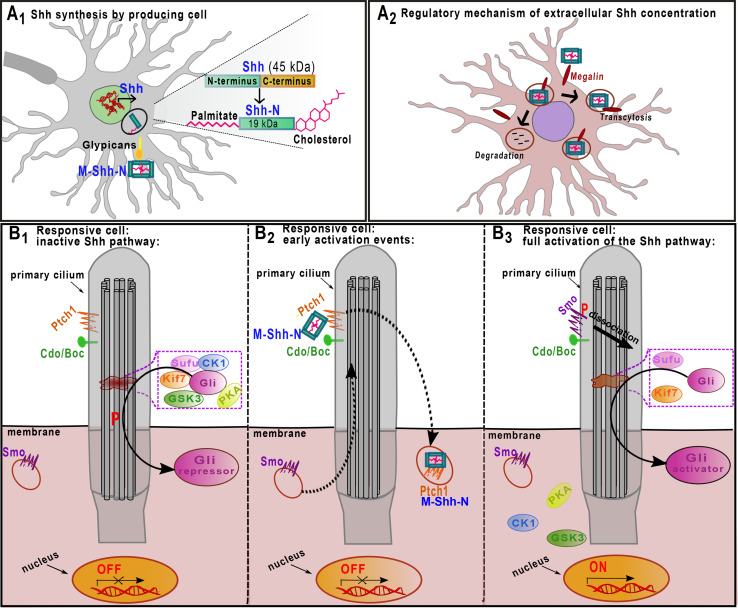
Shh signaling pathway in vertebrates. A1 Shh is translated as a 45-kDa precursor protein (Shh) and becomes an active signaling protein (Shh-N) after the addition of a cholesterol and a palmitate lipophilic moiety at its N-terminus. Shh is then trafficked to the cell surface and released from cells as a multimer (M-Shh-N). A2 Interactions with both megalin and glypicans regulate long-range Shh signaling. Binding to the transmembrane protein megalin promotes Shh internalization, resulting in either degradation or subsequent exocytosis. The Shh affinity for megalin is increased by glypicans. B1 Primary cilia are key organelles at which Shh signaling takes place. On their membranes, both receptors (Ptch1) and co-receptors (Cdo/Boc) are exposed. These structures also contain regulatory microtubule-associated complexes composed by Sufu, Kif7, PKA, GSK3, CK1, and Gli. In the absence of Shh, Ptch1 prevents membrane localization and activation of Smo, retaining it on intracellular vesicles. In this context, Gli2 and 3 (simply referred as Gli) proteins are held in the microtubule-associated complex, which induces their phosphorylation (red P). Upon phosphorylation, Gli leaves the complex and reaches the cytoplasm as a transcriptional repressor form (Gli repressor). B2 When Shh interacts with Ptch1 and Cdo/Boc, Smo is shuttled from an endocytic vesicle to the cilium, while the hedgehog–patched complex is internalized and degraded by lysosomes. B3 The de-repression of Smo, together with its phosphorylation, induces the dissociation of the Sufu–Gli-kinases complex, promoting the formation of the Gli activator form (Gli activator), which, after nuclear translocation, activates transcription of downstream targets
Fig. 2
Shh functions during cerebellar development. …
Fig. 2
Shh functions during cerebellar development. a During embryonic development, Shh is first secreted…
Fig. 2
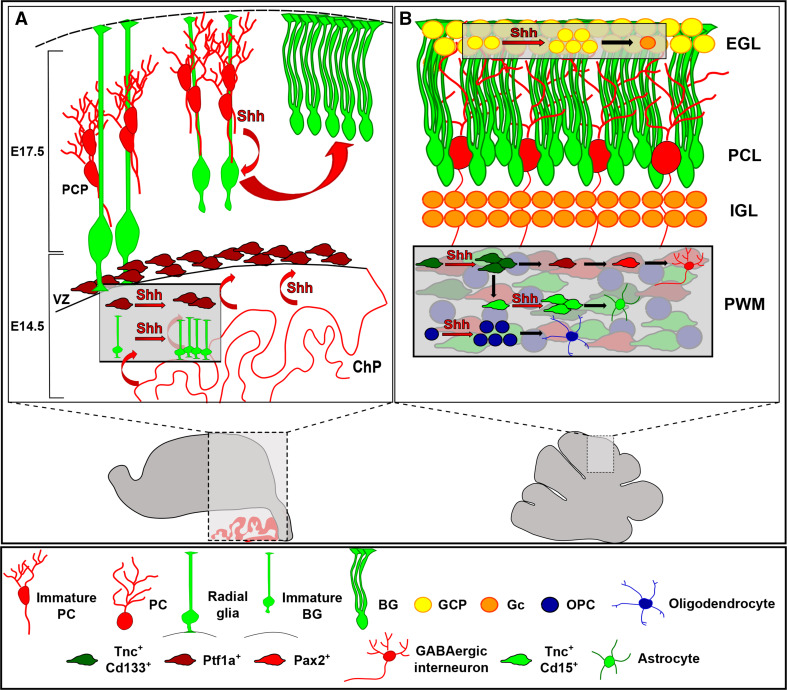
Shh functions during cerebellar development. a During embryonic development, Shh is first secreted by the choroid plexus (ChP) and is essential to radial glial cell proliferation and the expansion of Ptf1a+ progenitors of GABAergic neurons. Purkinje cells start Shh secretion by E17.5, modulating the correct differentiation of glial progenitors into mature BG. b Postnatally, Shh acts as a mitogen on both granule and oligodendrocyte precursor cells (GCPs and OPCs), in the EGL and PWM, respectively. In the PWM, Shh also exerts a proliferative function on the neural stem cell-like progenitors (Tnc+CD133+) that generate both intermediate astrocyte precursors (Tnc+CD15+) and GABAergic transient amplifying cells (Ptf1a+). PCP Purkinje cell progenitor, ChP choroid plexus, PWM prospective white matter, GL granular layer, PCL Purkinje cell layer, EGL external granular layer
Simpson F, Kerr MC, Wicking C. Trafficking, development and hedgehog. Mech Dev. 2009;126(5–6):279–288. doi: 10.1016/j.mod.2009.01.007.
-
DOI
-
PubMed
Tashiro S, Michiue T, Higashijima S, Zenno S, Ishimaru S, Takahashi F, Orihara M, Kojima T, Saigo K. Structure and expression of hedgehog, a Drosophila segment-polarity gene required for cell–cell communication. Gene. 1993;124(2):183–189. doi: 10.1016/0378-1119(93)90392-G.
-
DOI
-
PubMed
Nüsslein-Volhard C, Wieschaus E. Mutations affecting segment number and polarity in Drosophila . Nature. 1980;287(5785):795–801. doi: 10.1038/287795a0.
-
DOI
-
PubMed
Martinez-Arias A, Lawrence PA. Parasegments and compartments in the Drosophila embryo. Nature. 1985;313(6004):639–642. doi: 10.1038/313639a0.
-
DOI
-
PubMed
Wada H, Makabe K. Genome duplications of early vertebrates as a possible chronicle of the evolutionary history of the neural crest. Int J Biol Sci. 2006;2(3):133–141. doi: 10.7150/ijbs.2.133.
-
DOI
-
PMC
-
PubMed