

Chloroplast signaling within, between and beyond cells
- PMID: 26500659
- PMCID: PMC4593955
- DOI: 10.3389/fpls.2015.00781
Chloroplast signaling within, between and beyond cells
Abstract
The most conspicuous function of plastids is the oxygenic photosynthesis of chloroplasts, yet plastids are super-factories that produce a plethora of compounds that are indispensable for proper plant physiology and development. Given their origins as free-living prokaryotes, it is not surprising that plastids possess their own genomes whose expression is essential to plastid function. This semi-autonomous character of plastids requires the existence of sophisticated regulatory mechanisms that provide reliable communication between them and other cellular compartments. Such intracellular signaling is necessary for coordinating whole-cell responses to constantly varying environmental cues and cellular metabolic needs. This is achieved by plastids acting as receivers and transmitters of specific signals that coordinate expression of the nuclear and plastid genomes according to particular needs. In this review we will consider the so-called retrograde signaling occurring between plastids and nuclei, and between plastids and other organelles. Another important role of the plastid we will discuss is the involvement of plastid signaling in biotic and abiotic stress that, in addition to influencing retrograde signaling, has direct effects on several cellular compartments including the cell wall. We will also review recent evidence pointing to an intriguing function of chloroplasts in regulating intercellular symplasmic transport. Finally, we consider an intriguing yet less widely known aspect of plant biology, chloroplast signaling from the perspective of the entire plant. Thus, accumulating evidence highlights that chloroplasts, with their complex signaling pathways, provide a mechanism for exquisite regulation of plant development, metabolism and responses to the environment. As chloroplast processes are targeted for engineering for improved productivity the effect of such modifications on chloroplast signaling will have to be carefully considered in order to avoid unintended consequences on plant growth and development.
Keywords: cell wall; phytohormones; plasmodesmata; plastid signaling; redox; retrograde signaling; stress responses; stromules.
Figures
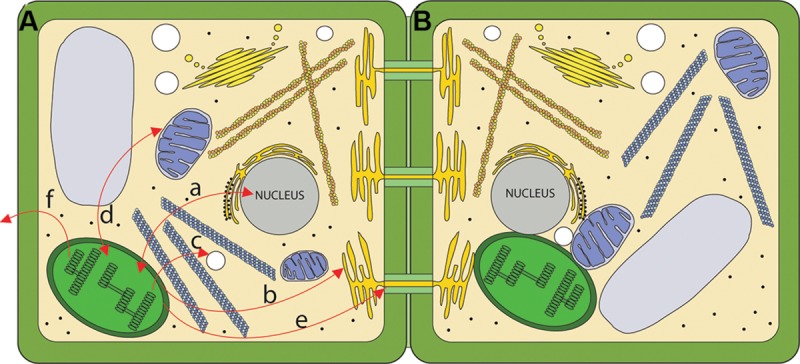
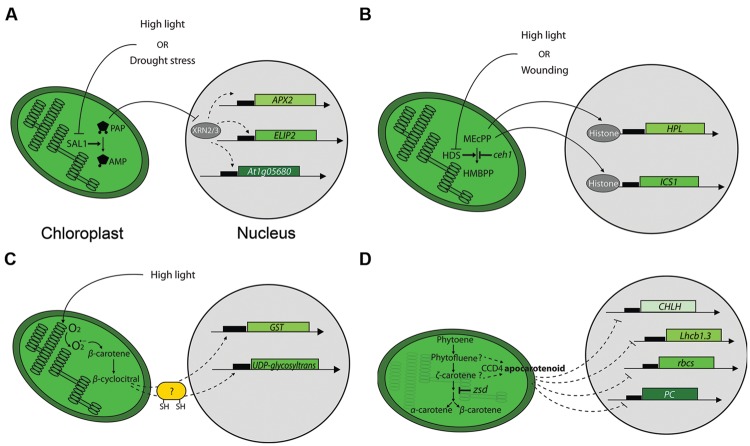
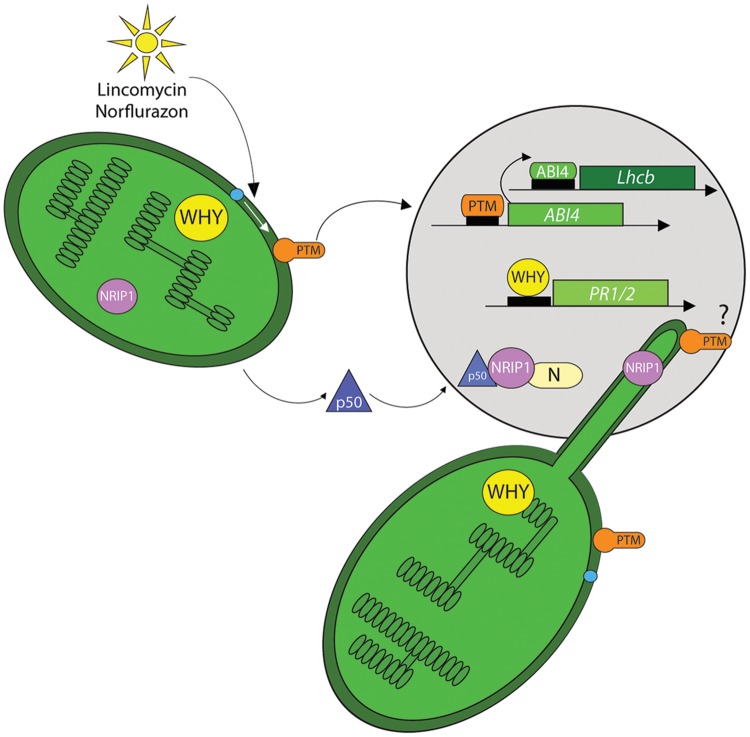
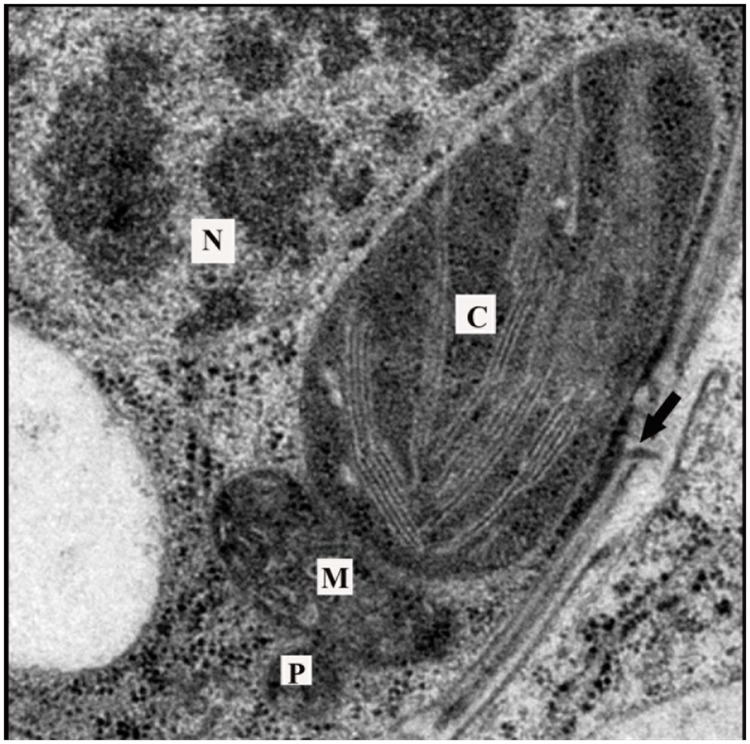
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials