

TIMP3 interplays with apelin to regulate cardiovascular metabolism in hypercholesterolemic mice
- PMID: 26500845
- PMCID: PMC4588459
- DOI: 10.1016/j.molmet.2015.07.007
TIMP3 interplays with apelin to regulate cardiovascular metabolism in hypercholesterolemic mice
Abstract
Objective: Tissue inhibitor of metalloproteinase 3 (TIMP3) is an extracellular matrix (ECM) bound protein, which has been shown to be downregulated in human subjects and experimental models with cardiometabolic disorders, including type 2 diabetes mellitus, hypertension and atherosclerosis. The aim of this study was to investigate the effects of TIMP3 on cardiac energy homeostasis during increased metabolic stress conditions.
Methods: ApoE(-/-)TIMP3(-/-) and ApoE(-/-) mice on a C57BL/6 background were subjected to telemetric ECG analysis and experimental myocardial infarction as models of cardiac stress induction. We used Western blot, qRT-PCR, histology, metabolomics, RNA-sequencing and in vivo phenotypical analysis to investigate the molecular mechanisms of altered cardiac energy metabolism.
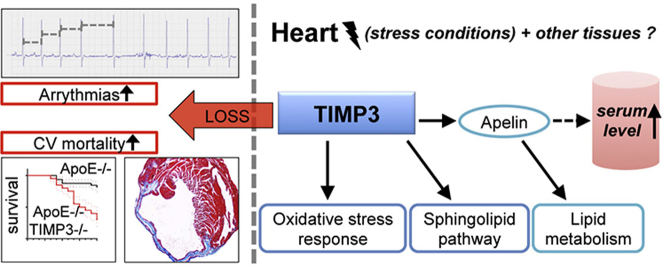
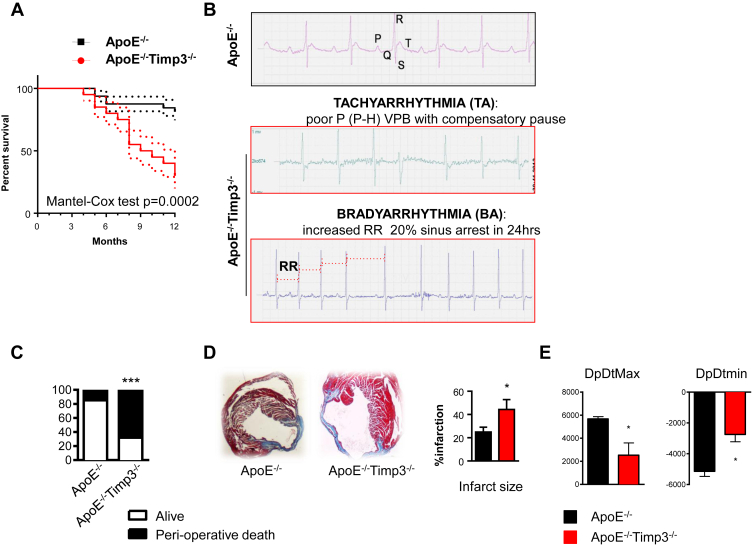
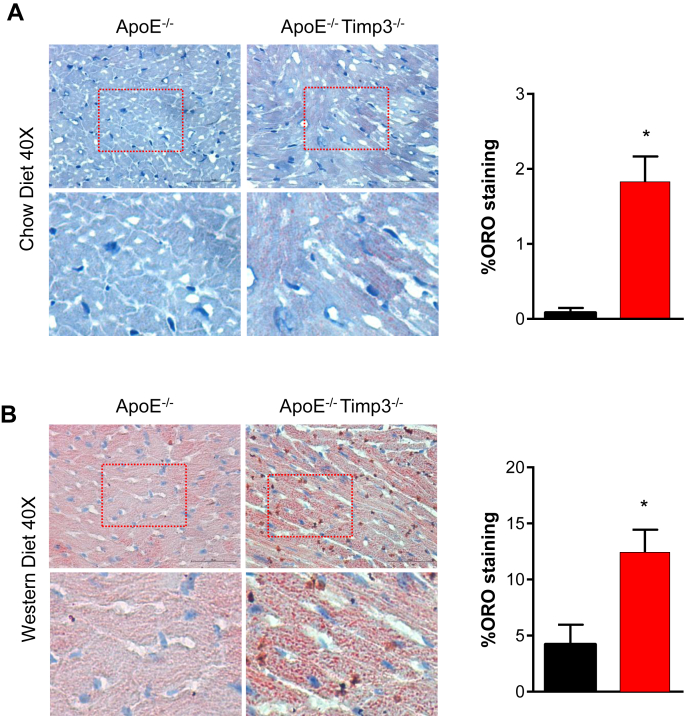
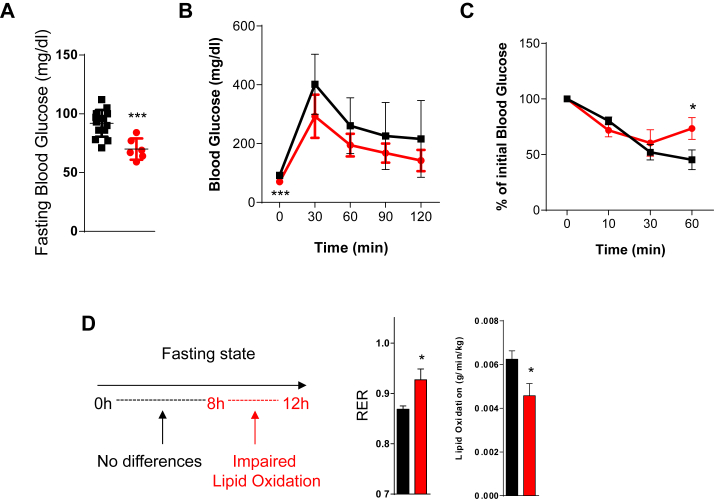
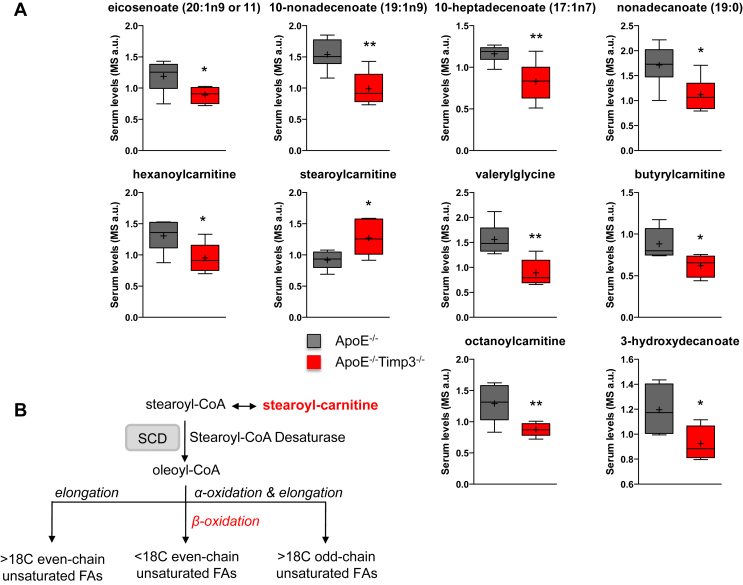
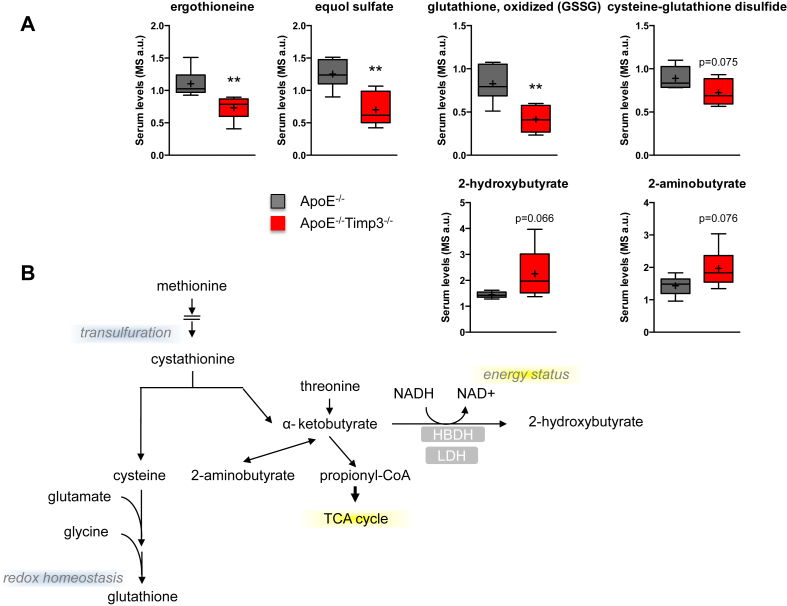
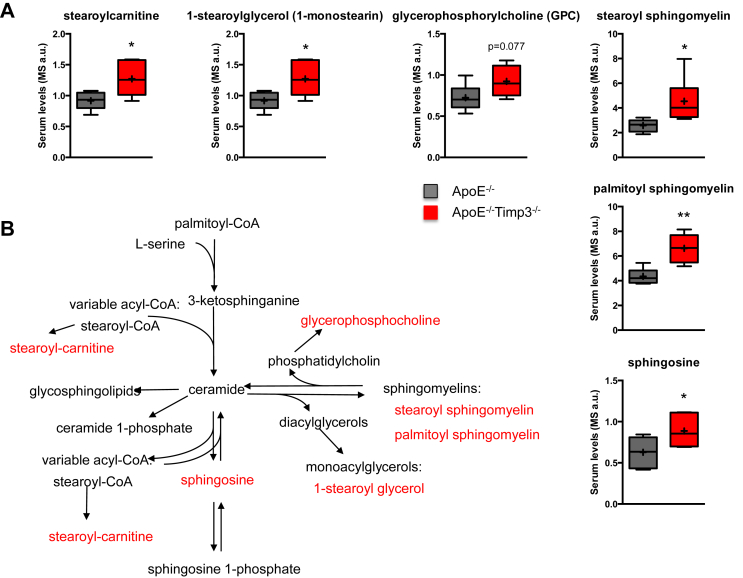
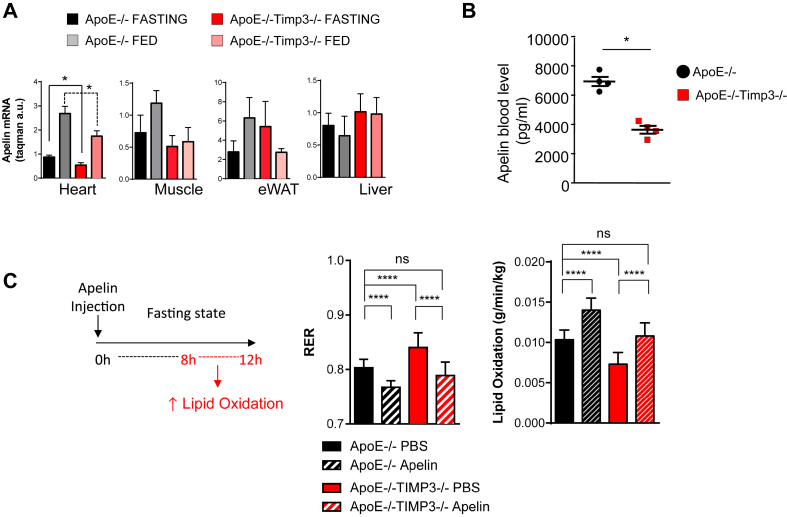
Results: ApoE(-/-)TIMP3(-/-) revealed decreased lifespan. Telemetric ECG analysis showed increased arrhythmic episodes, and experimental myocardial infarction by left anterior descending artery (LAD) ligation resulted in increased peri-operative mortality together with increased scar formation, ventricular dilatation and a reduction of cardiac function after 4 weeks in the few survivors. Hearts of ApoE(-/-)TIMP3(-/-) exhibited accumulation of neutral lipids when fed a chow diet, which was exacerbated by a high fat, high cholesterol diet. Metabolomics analysis revealed an increase in circulating markers of oxidative stress with a reduction in long chain fatty acids. Using whole heart mRNA sequencing, we identified apelin as a putative modulator of these metabolic defects. Apelin is a regulator of fatty acid oxidation, and we found a reduction in the levels of enzymes involved in fatty acid oxidation in the left ventricle of ApoE(-/-)TIMP3(-/-) mice. Injection of apelin restored the hitherto identified metabolic defects of lipid oxidation.
Conclusion: TIMP3 regulates lipid metabolism as well as oxidative stress response via apelin. These findings therefore suggest that TIMP3 maintains metabolic flexibility in the heart, particularly during episodes of increased cardiac stress.
Keywords: Apelin; Arrhythmia; Heart; Lipid metabolism; Oxidative stress; TIMP3.
Figures
References
-
- Lopaschuk G.D., Ussher J.R., Folmes C.D., Jaswal J.S., Stanley W.C. Myocardial fatty acid metabolism in health and disease. Physiological Reviews. 2010;90:207–258. - PubMed
-
- Carnevale D., Lembo G. Placental growth factor and cardiac inflammation. Trends in Cardiovascular Medicine. 2012;22:209–212. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
