

Metazoan Hsp70-based protein disaggregases: emergence and mechanisms
- PMID: 26501065
- PMCID: PMC4598581
- DOI: 10.3389/fmolb.2015.00057
Metazoan Hsp70-based protein disaggregases: emergence and mechanisms
Abstract
Proteotoxic stresses and aging cause breakdown of cellular protein homeostasis, allowing misfolded proteins to form aggregates, which dedicated molecular machines have evolved to solubilize. In bacteria, fungi, protozoa and plants protein disaggregation involves an Hsp70•J-protein chaperone system, which loads and activates a powerful AAA+ ATPase (Hsp100) disaggregase onto protein aggregate substrates. Metazoans lack cytosolic and nuclear Hsp100 disaggregases but still eliminate protein aggregates. This longstanding puzzle of protein quality control is now resolved. Robust protein disaggregation activity recently shown for the metazoan Hsp70-based disaggregases relies instead on a crucial cooperation between two J-protein classes and interaction with the Hsp110 co-chaperone. An expanding multiplicity of Hsp70 and J-protein family members in metazoan cells facilitates different configurations of this Hsp70-based disaggregase allowing unprecedented versatility and specificity in protein disaggregation. Here we review the architecture, operation, and adaptability of the emerging metazoan disaggregation system and discuss how this evolved.
Keywords: Hsp110; Hsp70; J-protein; metazoan; protein disaggregation.
Figures
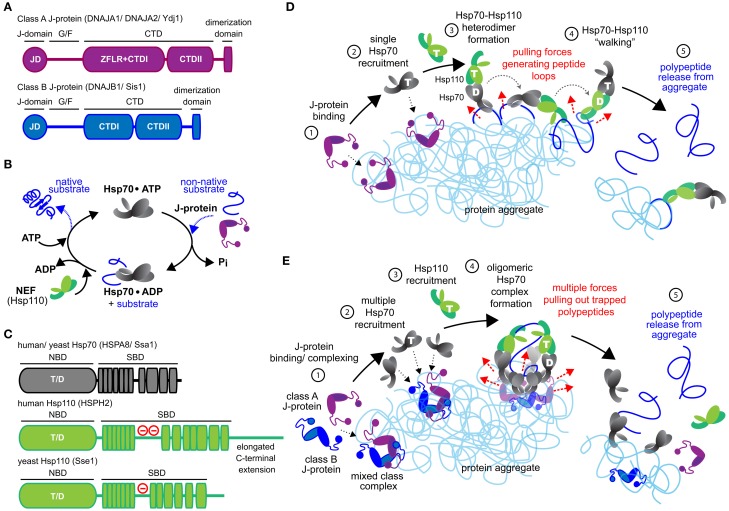
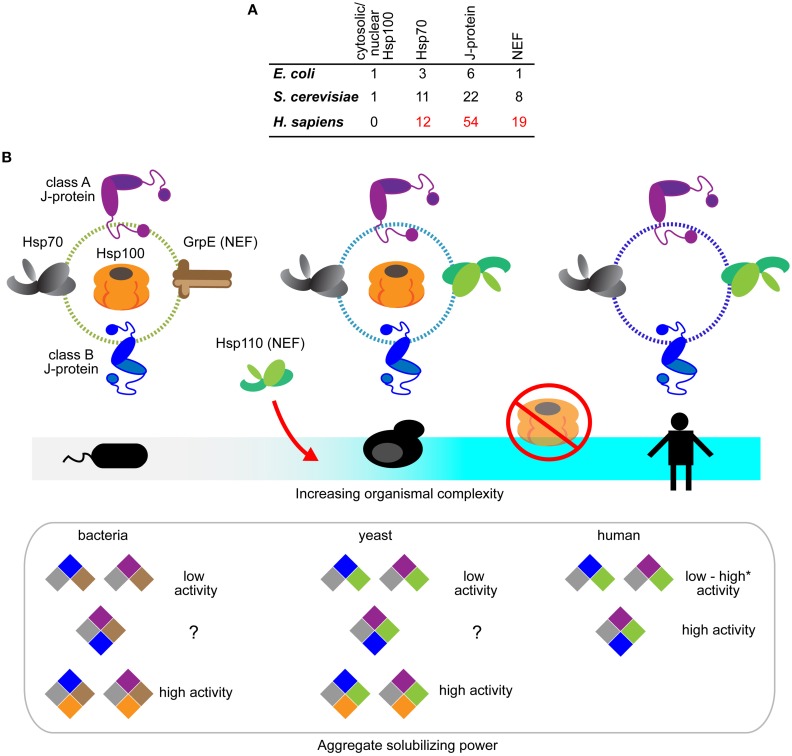
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous