

Macromolecular crowding creates heterogeneous environments of gene expression in picolitre droplets
- PMID: 26501750
- PMCID: PMC4740931
- DOI: 10.1038/nnano.2015.243
Macromolecular crowding creates heterogeneous environments of gene expression in picolitre droplets
Abstract
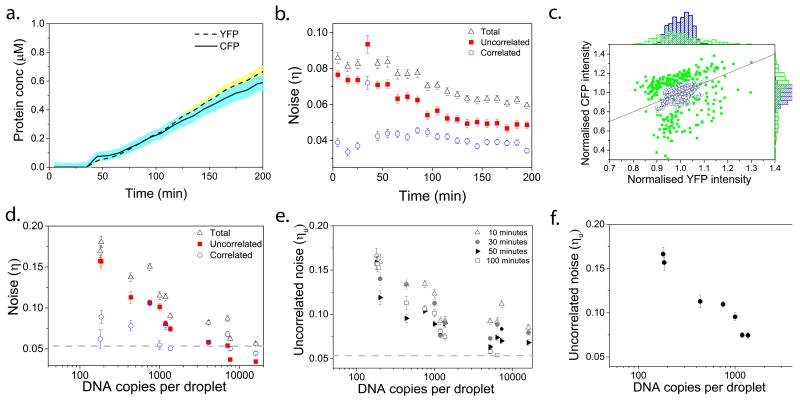
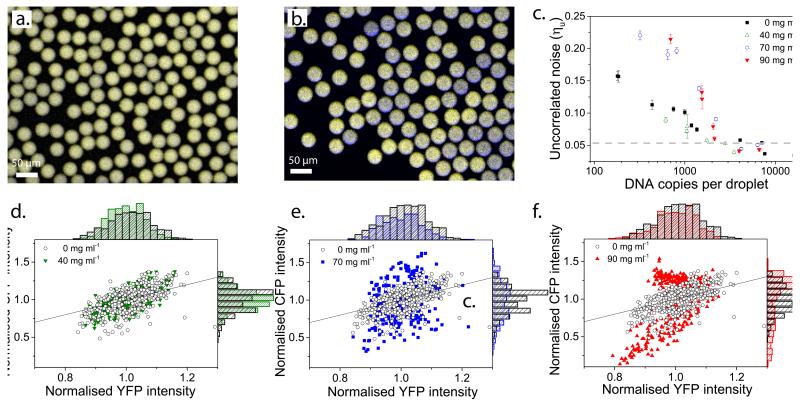
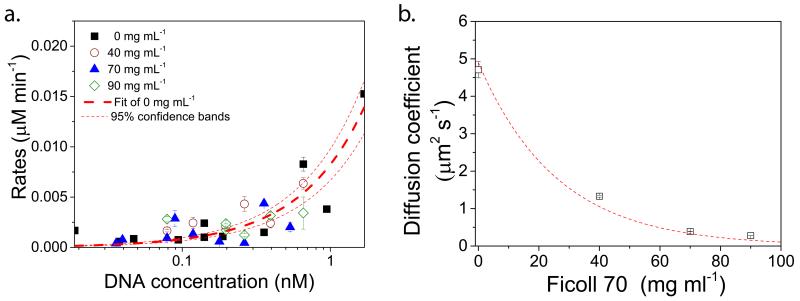
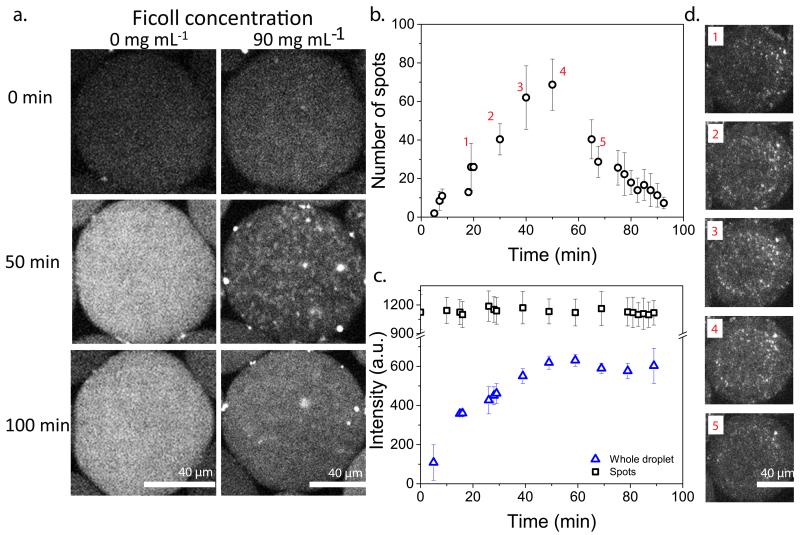
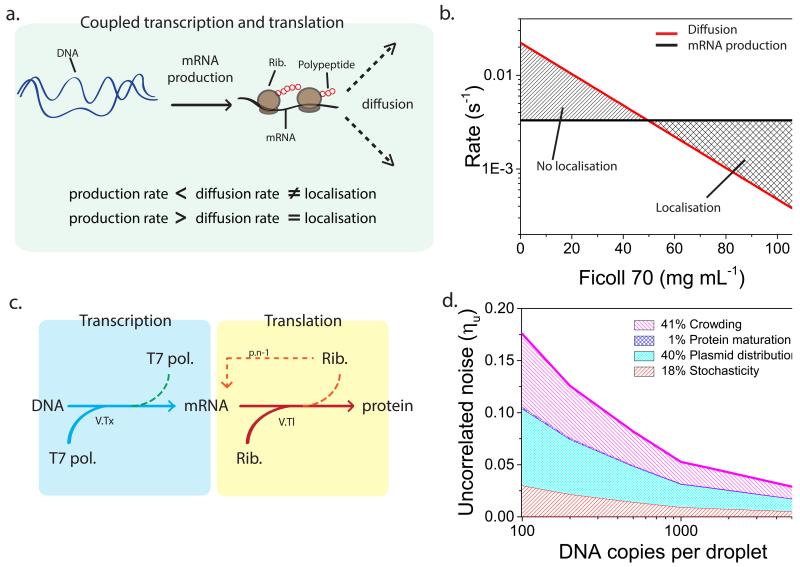
Understanding the dynamics of complex enzymatic reactions in highly crowded small volumes is crucial for the development of synthetic minimal cells. Compartmentalized biochemical reactions in cell-sized containers exhibit a degree of randomness due to the small number of molecules involved. However, it is unknown how the physical environment contributes to the stochastic nature of multistep enzymatic processes. Here, we present a robust method to quantify gene expression noise in vitro using droplet microfluidics. We study the changes in stochasticity in the cell-free gene expression of two genes compartmentalized within droplets as a function of DNA copy number and macromolecular crowding. We find that decreased diffusion caused by a crowded environment leads to the spontaneous formation of heterogeneous microenvironments of mRNA as local production rates exceed the diffusion rates of macromolecules. This heterogeneity leads to a higher probability of the molecular machinery staying in the same microenvironment, directly increasing the system's stochasticity.
Figures

References
-
- Graf T, Stadtfeld M. Heterogeneity of Embryonic and Adult Stem Cells. Cell Stem Cell. 2008;3:480–483. - PubMed
-
- Gupta Piyush B., et al. Stochastic State Transitions Give Rise to Phenotypic Equilibrium in Populations of Cancer Cells. Cell. 2011;146:633–644. - PubMed
-
- Weinberger LS, Burnett JC, Toettcher JE, Arkin AP, Schaffer DV. Stochastic Gene Expression in a Lentiviral Positive-Feedback Loop: HIV-1 Tat Fluctuations Drive Phenotypic Diversity. Cell. 2005;122:169–182. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
