

In vivo characterization of the physicochemical properties of polymer-linked TLR agonists that enhance vaccine immunogenicity
- PMID: 26501954
- PMCID: PMC5842712
- DOI: 10.1038/nbt.3371
In vivo characterization of the physicochemical properties of polymer-linked TLR agonists that enhance vaccine immunogenicity
Abstract
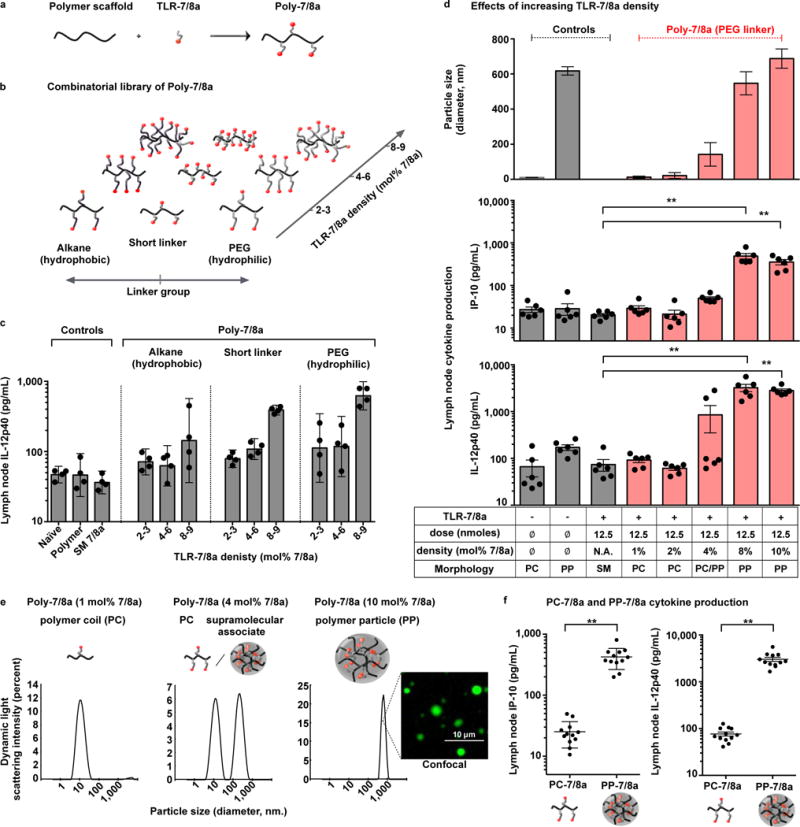
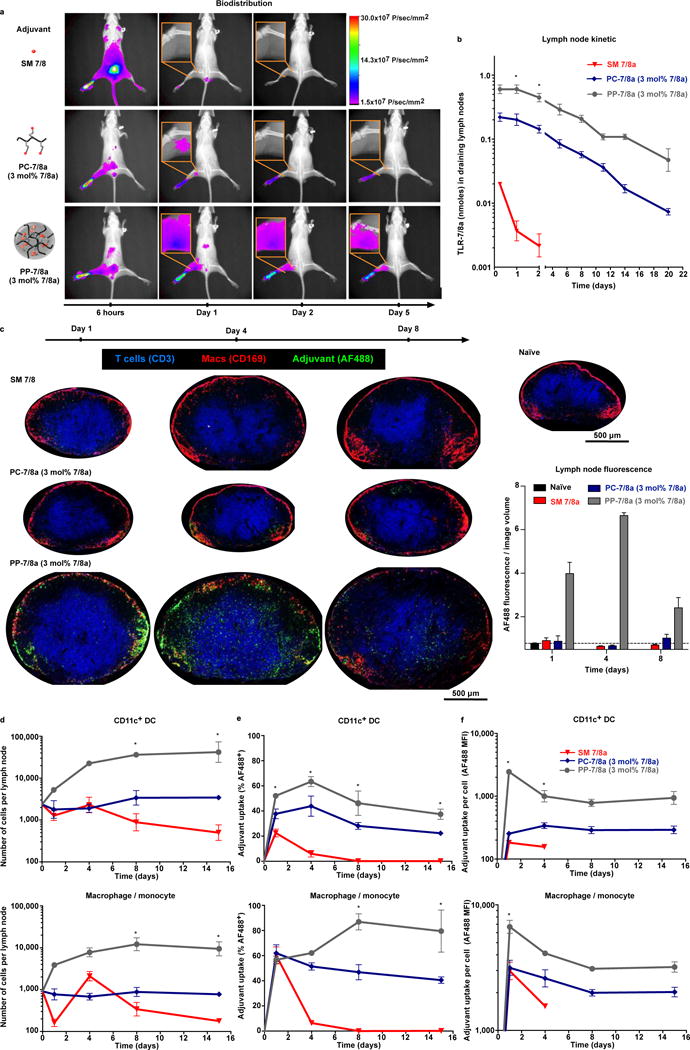
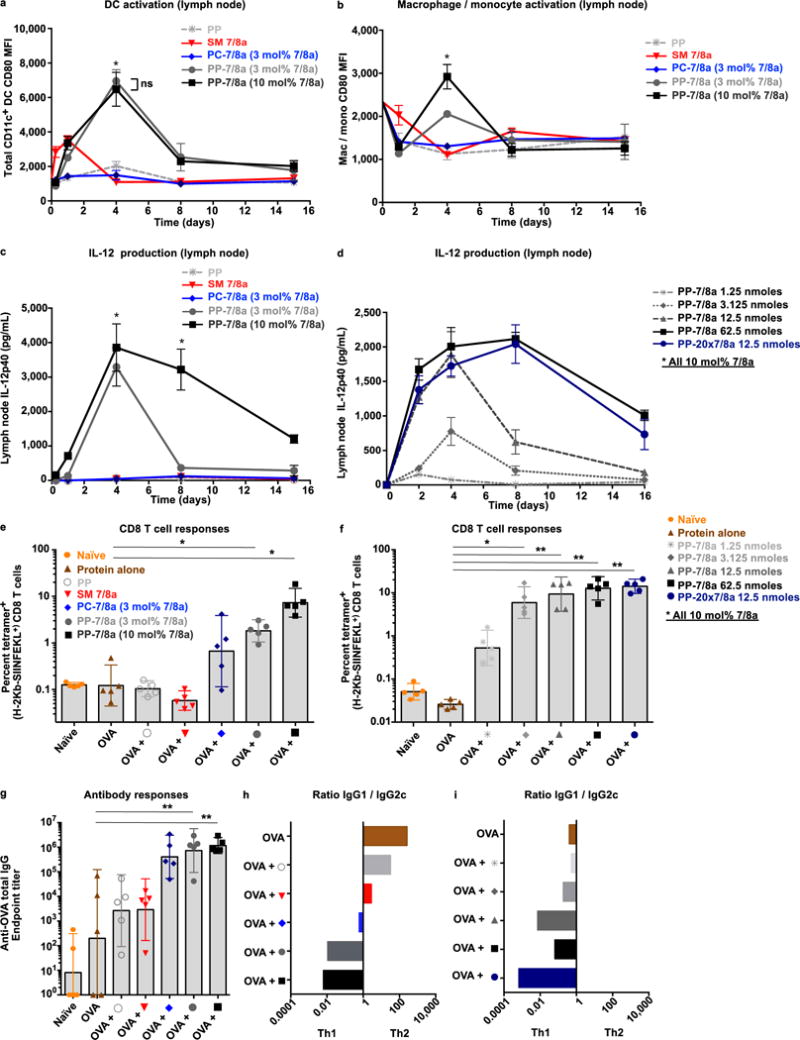
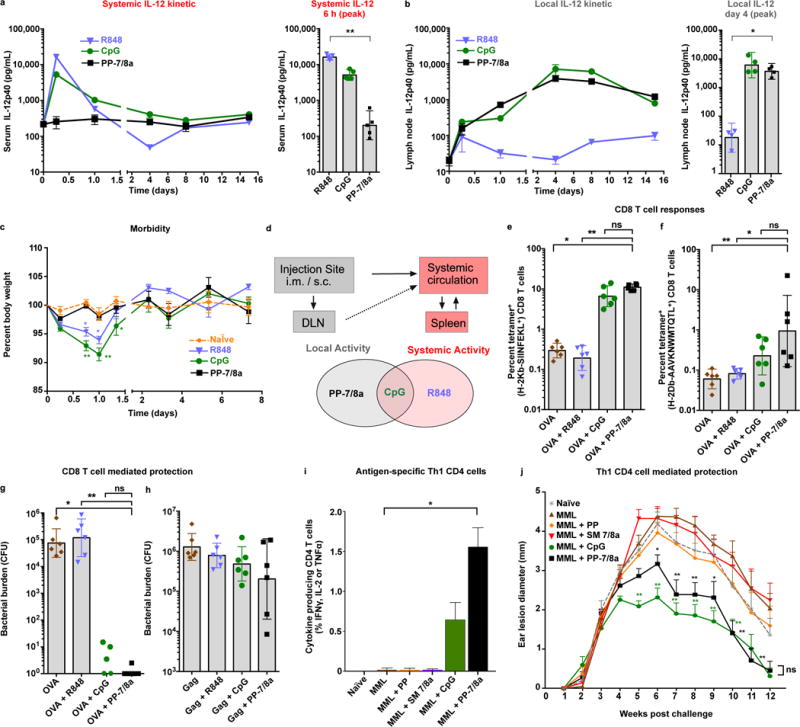
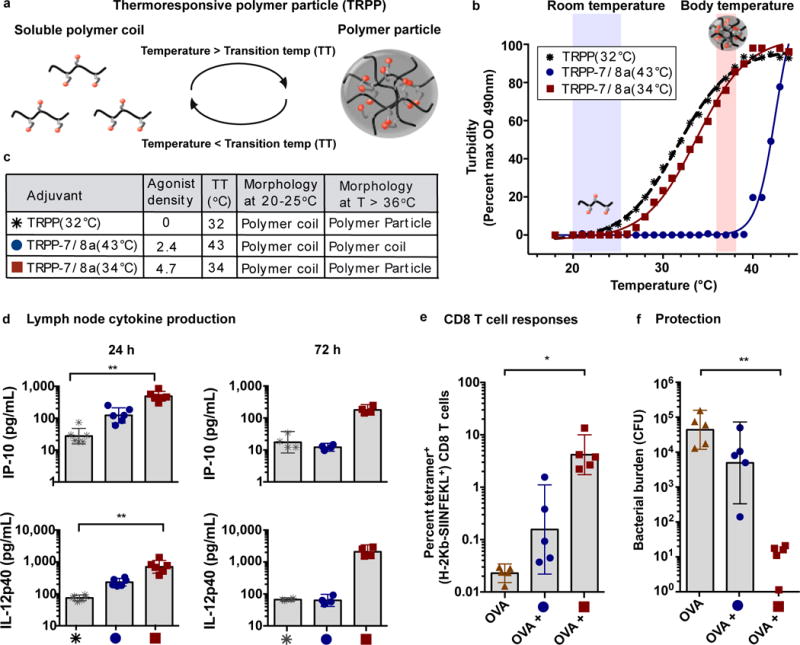
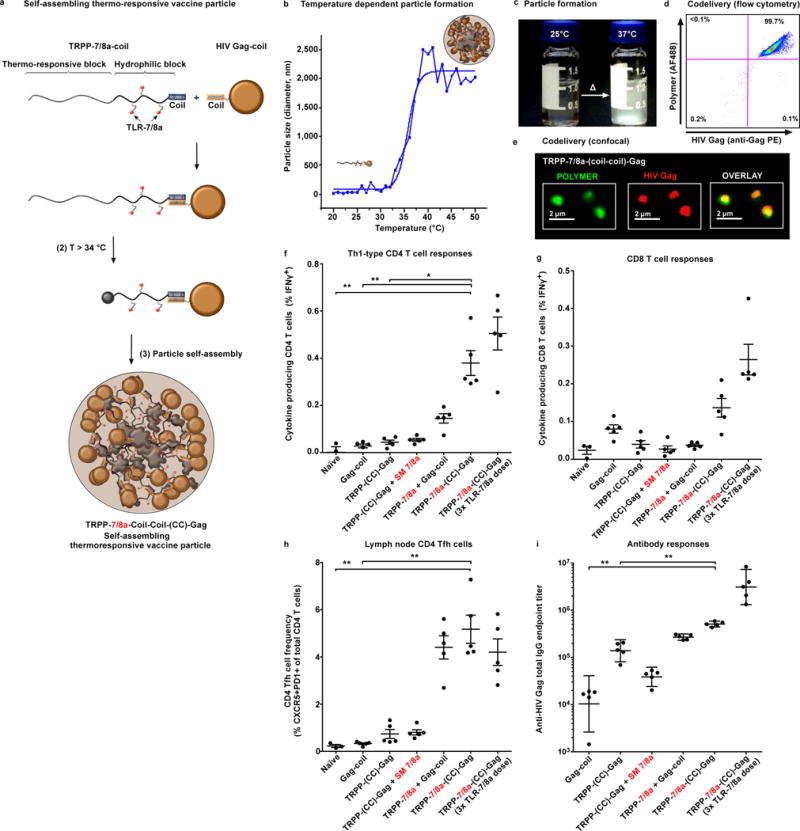
The efficacy of vaccine adjuvants such as Toll-like receptor agonists (TLRa) can be improved through formulation and delivery approaches. Here, we attached small molecule TLR-7/8a to polymer scaffolds (polymer-TLR-7/8a) and evaluated how different physicochemical properties of the TLR-7/8a and polymer carrier influenced the location, magnitude and duration of innate immune activation in vivo. Particle formation by polymer-TLR-7/8a was the most important factor for restricting adjuvant distribution and prolonging activity in draining lymph nodes. The improved pharmacokinetic profile by particulate polymer-TLR-7/8a was also associated with reduced morbidity and enhanced vaccine immunogenicity for inducing antibodies and T cell immunity. We extended these findings to the development of a modular approach in which protein antigens are site-specifically linked to temperature-responsive polymer-TLR-7/8a adjuvants that self-assemble into immunogenic particles at physiologic temperatures in vivo. Our findings provide a chemical and structural basis for optimizing adjuvant design to elicit broad-based antibody and T cell responses with protein antigens.
Conflict of interest statement
G.M.L., R.L., K.D.F, L.W.S. and R.A.S. are listed as inventors on patents describing polymer-based vaccines. K.D.F and L.W.S. are scientific founders and equity holders in PsiOxus Therapeutics, Ltd. (Oxford, UK). G.M.L. and J.R.F. are scientific founders and equity holders in Avidea Technologies, Inc., which is developing polymer-based technologies for immunotherapeutic applications (Baltimore, MD, USA).
Figures
Comment in
-
Big thinking for adjuvants.Nat Biotechnol. 2015 Nov;33(11):1146-8. doi: 10.1038/nbt.3398. Nat Biotechnol. 2015. PMID: 26544144 No abstract available.
References
-
- Epstein JE, et al. Live attenuated malaria vaccine designed to protect through hepatic CD8(+) T cell immunity. Science. 2011;334:475–480. - PubMed
-
- Andersen P, Woodworth JS. Tuberculosis vaccines–rethinking the current paradigm. Trends Immunol. 2014;35:387–395. - PubMed
-
- Arens R, van Hall T, van der Burg SH, Ossendorp F, Melief CJ. Prospects of combinatorial synthetic peptide vaccine-based immunotherapy against cancer. Seminars in immunology. 2013;25:182–190. - PubMed
-
- Guy B. The perfect mix: recent progress in adjuvant research. Nature reviews Microbiology. 2007;5:505–517. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
