

Ion channels enable electrical communication in bacterial communities
- PMID: 26503040
- PMCID: PMC4890463
- DOI: 10.1038/nature15709
Ion channels enable electrical communication in bacterial communities
Abstract
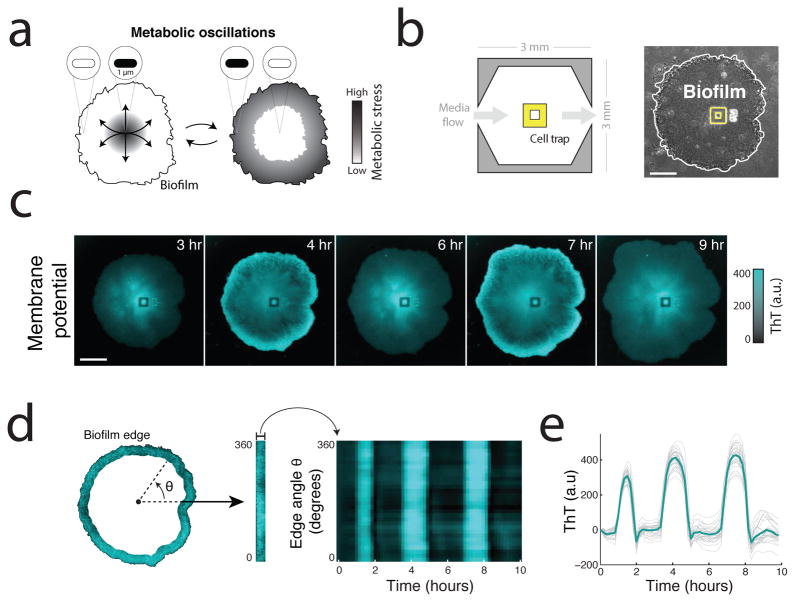
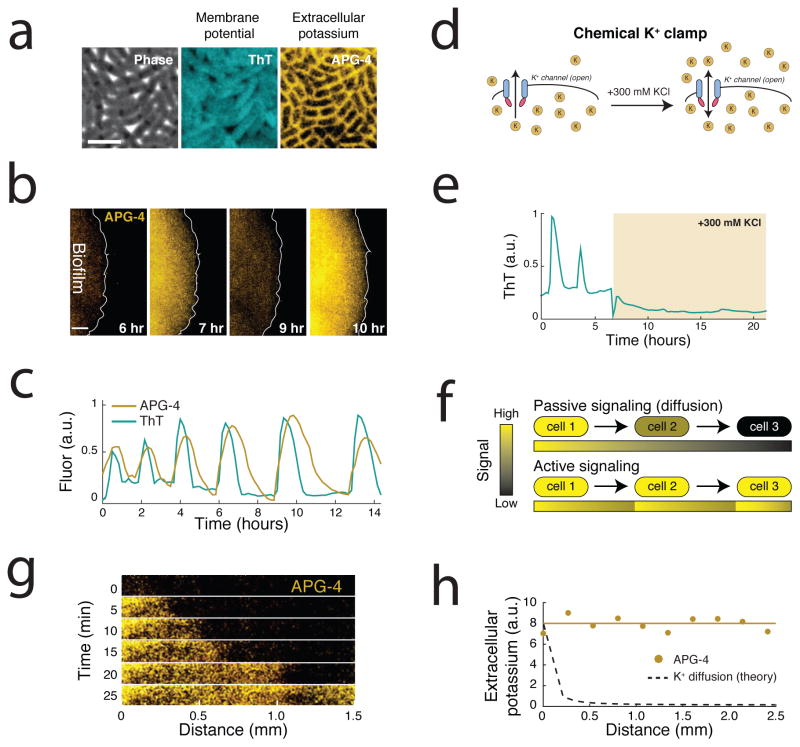
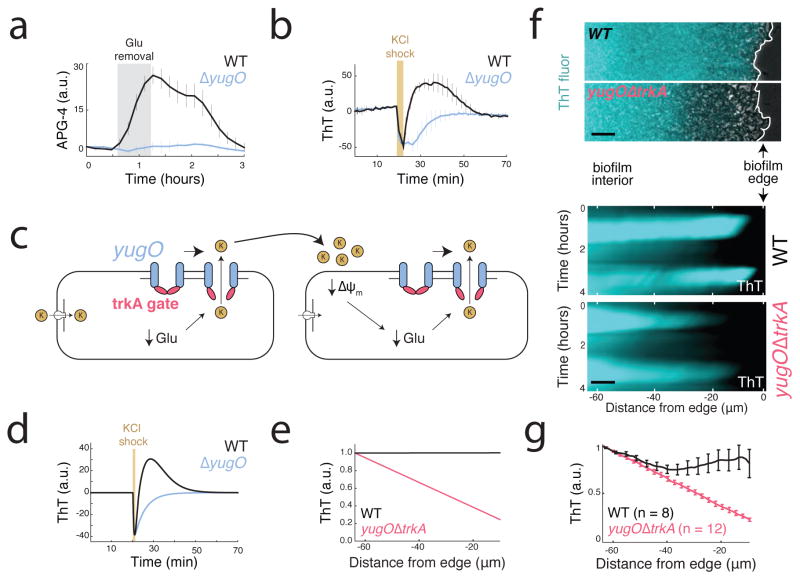
The study of bacterial ion channels has provided fundamental insights into the structural basis of neuronal signalling; however, the native role of ion channels in bacteria has remained elusive. Here we show that ion channels conduct long-range electrical signals within bacterial biofilm communities through spatially propagating waves of potassium. These waves result from a positive feedback loop, in which a metabolic trigger induces release of intracellular potassium, which in turn depolarizes neighbouring cells. Propagating through the biofilm, this wave of depolarization coordinates metabolic states among cells in the interior and periphery of the biofilm. Deletion of the potassium channel abolishes this response. As predicted by a mathematical model, we further show that spatial propagation can be hindered by specific genetic perturbations to potassium channel gating. Together, these results demonstrate a function for ion channels in bacterial biofilms, and provide a prokaryotic paradigm for active, long-range electrical signalling in cellular communities.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
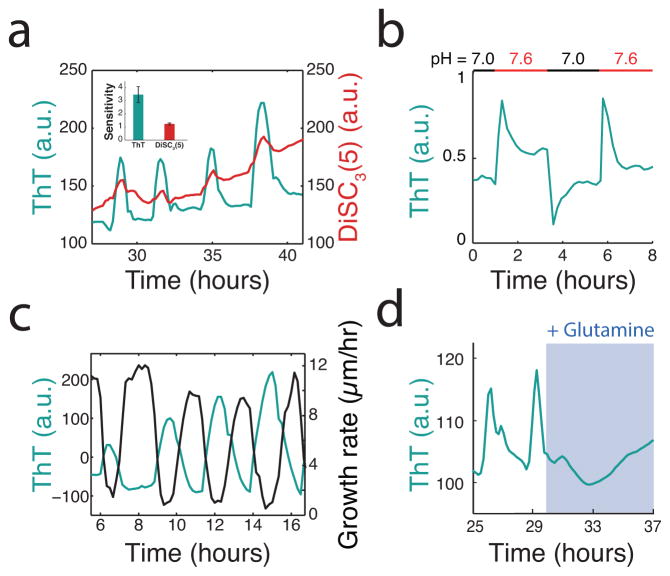
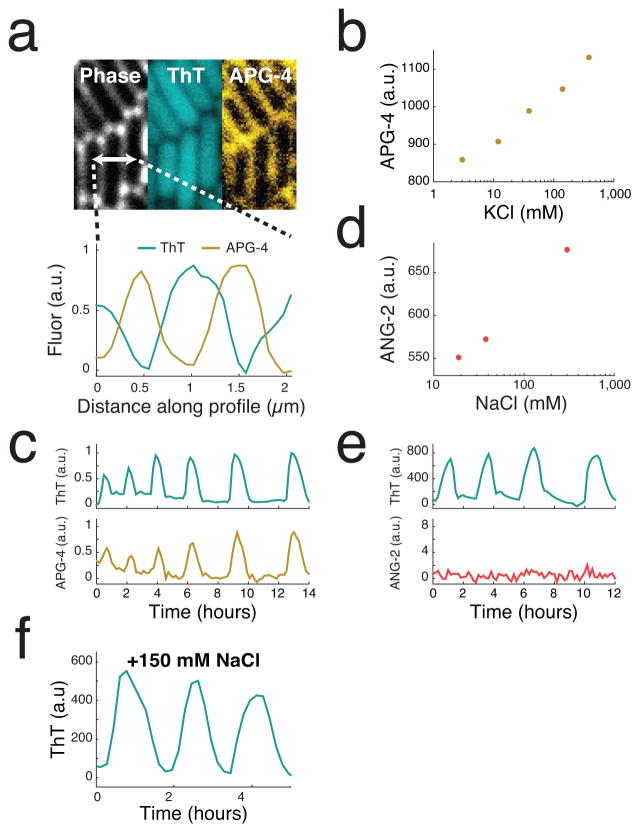
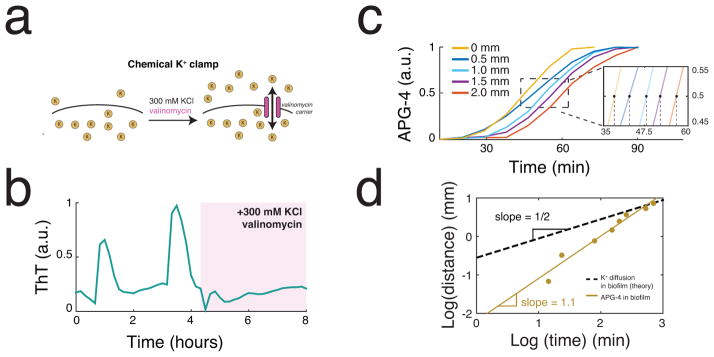
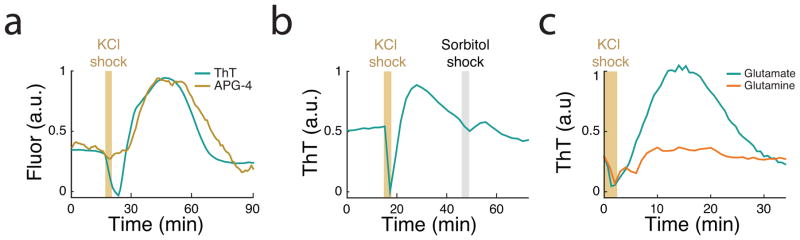
Comment in
-
Microbiology: Electrical signalling goes bacterial.Nature. 2015 Nov 5;527(7576):44-5. doi: 10.1038/nature15641. Epub 2015 Oct 21. Nature. 2015. PMID: 26503058 No abstract available.
-
Biofilms: Electrifying long-range signalling.Nat Rev Microbiol. 2015 Dec;13(12):737. doi: 10.1038/nrmicro3588. Epub 2015 Nov 9. Nat Rev Microbiol. 2015. PMID: 26548916 No abstract available.
References
-
- Hille B. Ion Channels of Excitable Membranes. 2001
-
- MacKinnon R. Potassium channels and the atomic basis of selective ion conduction. Bioscience Reports. 2004;24:75–100. - PubMed
-
- Doyle DA, et al. The structure of the potassium channel: molecular basis of K+ conduction and selectivity. Science. 1998;280:69–77. - PubMed
-
- Ren D, et al. A prokaryotic voltage-gated sodium channel. Science. 2001;294:2372–2375. - PubMed
-
- Iyer R, Iverson TM, Accardi A, Miller C. A biological role for prokaryotic ClC chloride channels. Nature. 2002;419:715–718. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
