

Disrupting astrocyte-neuron lactate transfer persistently reduces conditioned responses to cocaine
- PMID: 26503760
- PMCID: PMC4960452
- DOI: 10.1038/mp.2015.157
Disrupting astrocyte-neuron lactate transfer persistently reduces conditioned responses to cocaine
Abstract
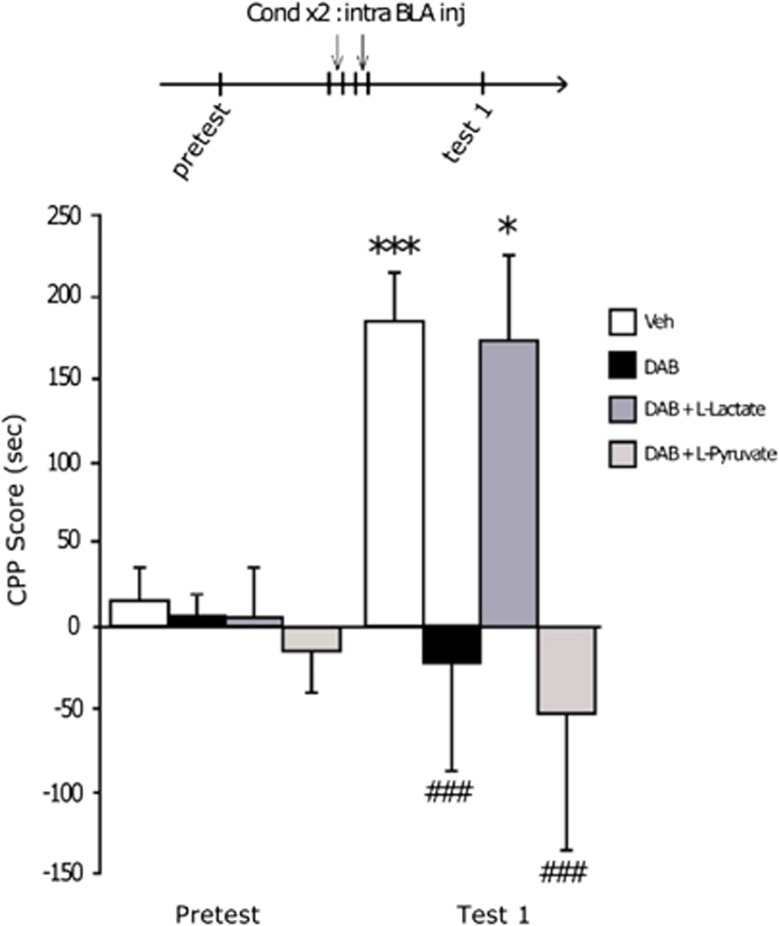
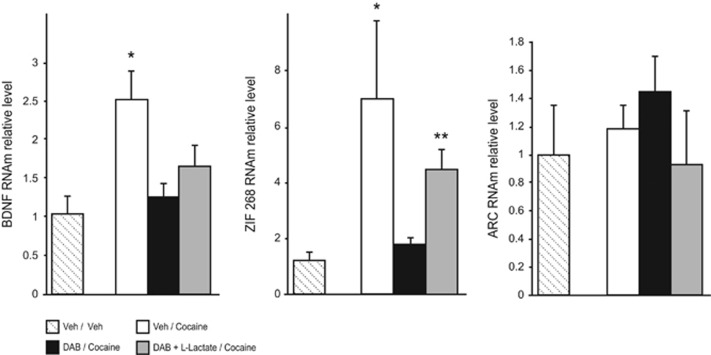
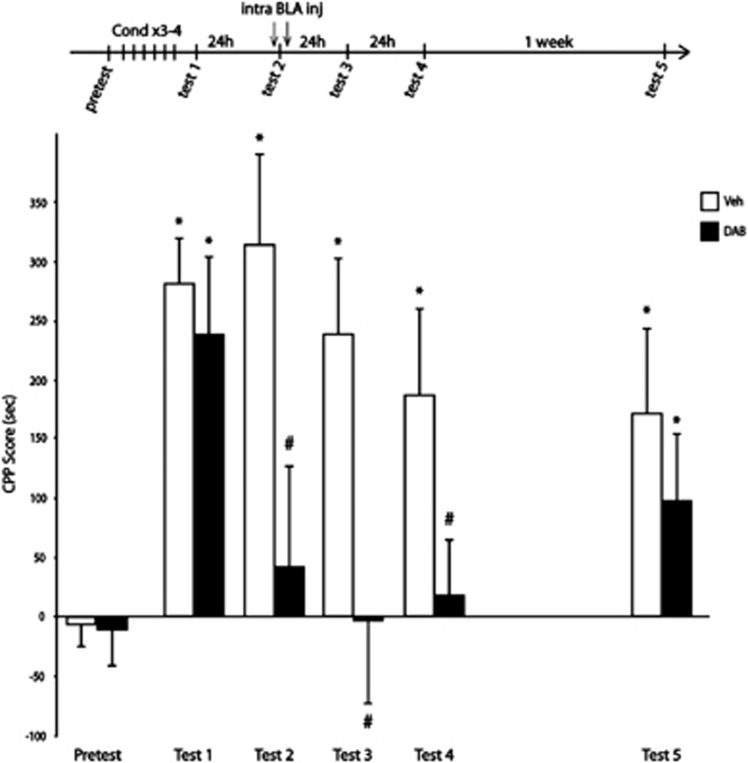
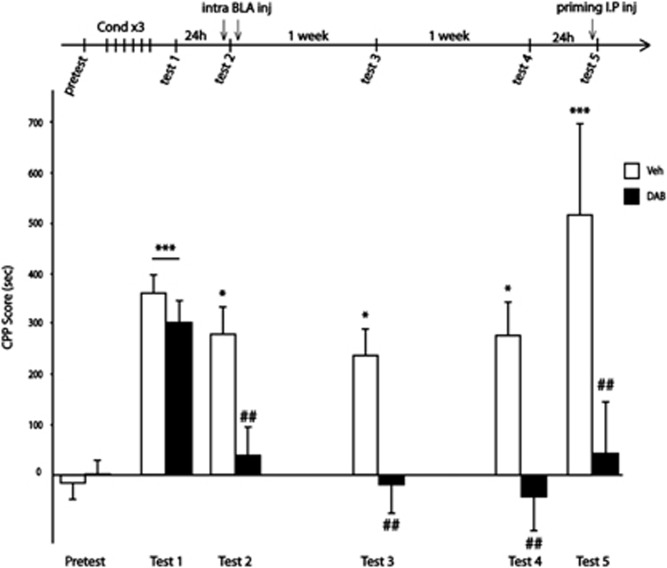
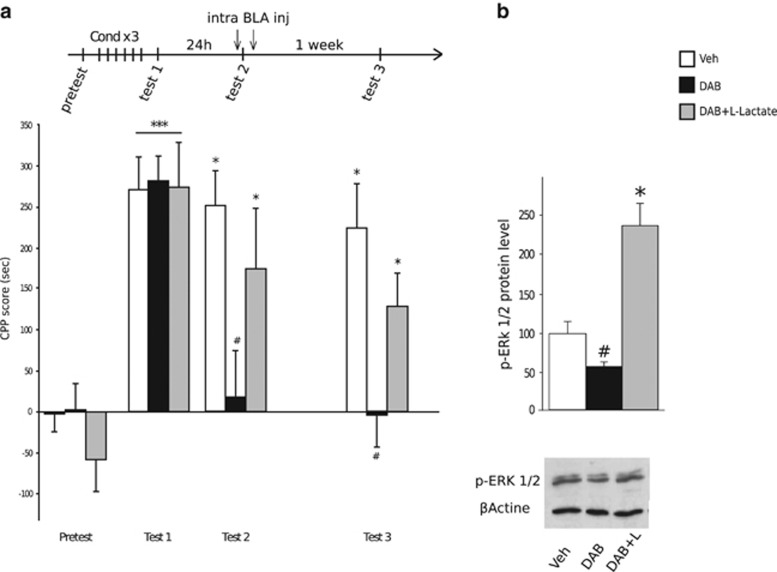
A central problem in the treatment of drug addiction is the high risk of relapse often precipitated by drug-associated cues. The transfer of glycogen-derived lactate from astrocytes to neurons is required for long-term memory. Whereas blockade of drug memory reconsolidation represents a potential therapeutic strategy, the role of astrocyte-neuron lactate transport in long-term conditioning has received little attention. By infusing an inhibitor of glycogen phosphorylase into the basolateral amygdala of rats, we report that disruption of astrocyte-derived lactate not only transiently impaired the acquisition of a cocaine-induced conditioned place preference but also persistently disrupted an established conditioning. The drug memory was rescued by L-Lactate co-administration through a mechanism requiring the synaptic plasticity-related transcription factor Zif268 and extracellular signal-regulated kinase (ERK) signalling pathway but not the brain-derived neurotrophic factor (Bdnf). The long-term amnesia induced by glycogenolysis inhibition and the concomitant decreased expression of phospho-ERK were both restored with L-Lactate co-administration. These findings reveal a critical role for astrocyte-derived lactate in positive memory formation and highlight a novel amygdala-dependent reconsolidation process, whose disruption may offer a novel therapeutic target to reduce the long-lasting conditioned responses to cocaine.
Figures
References
-
- Hyman SE. Addiction: a disease of learning and memory. Am J Psychiatry 2005; 162: 1414–1422. - PubMed
-
- O'Brien CP, Childress AR, McLellan AT, Ehrman R. Classical conditioning in drug-dependent humans. Ann NY Acad Sci 1992; 654: 400–415. - PubMed
-
- Childress AR, McLellan AT, Ehrman R, O'Brien CP. Classically conditioned responses in opioid and cocaine dependence: a role in relapse? NIDA Res Monogr 1988; 84: 25–43. - PubMed
-
- de Wit H, Stewart J. Reinstatement of cocaine-reinforced responding in the rat. Psychopharmacology 1981; 75: 134–143. - PubMed
-
- Meil WM, See RE. Conditioned cued recovery of responding following prolonged withdrawal from self-administered cocaine in rats: an animal model of relapse. Behav Pharmacol 1996; 7: 754–763. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
