

Sites of retroviral DNA integration: From basic research to clinical applications
- PMID: 26508664
- PMCID: PMC4866806
- DOI: 10.3109/10409238.2015.1102859
Sites of retroviral DNA integration: From basic research to clinical applications
Abstract
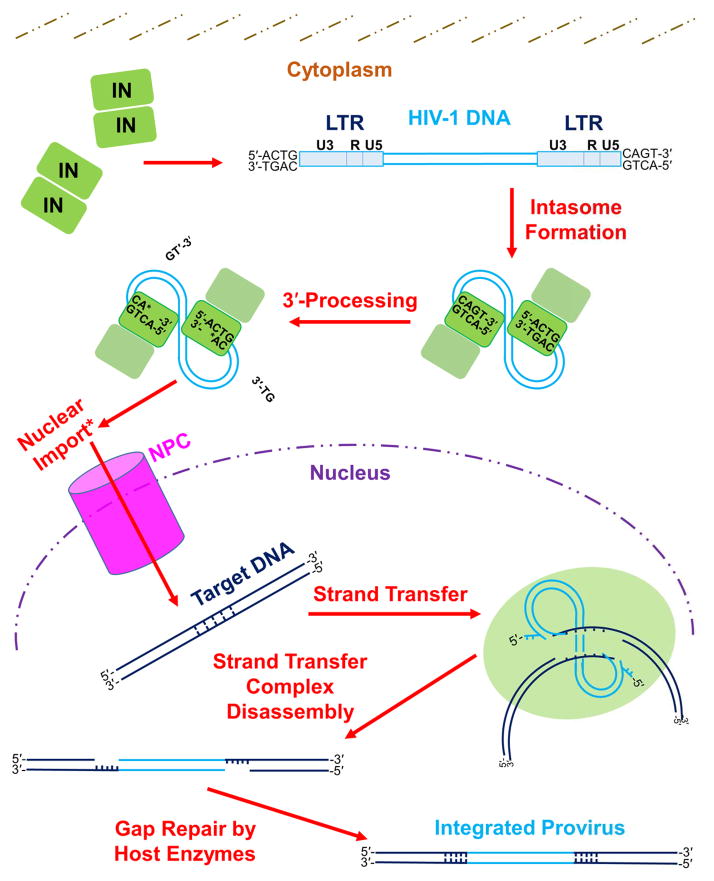
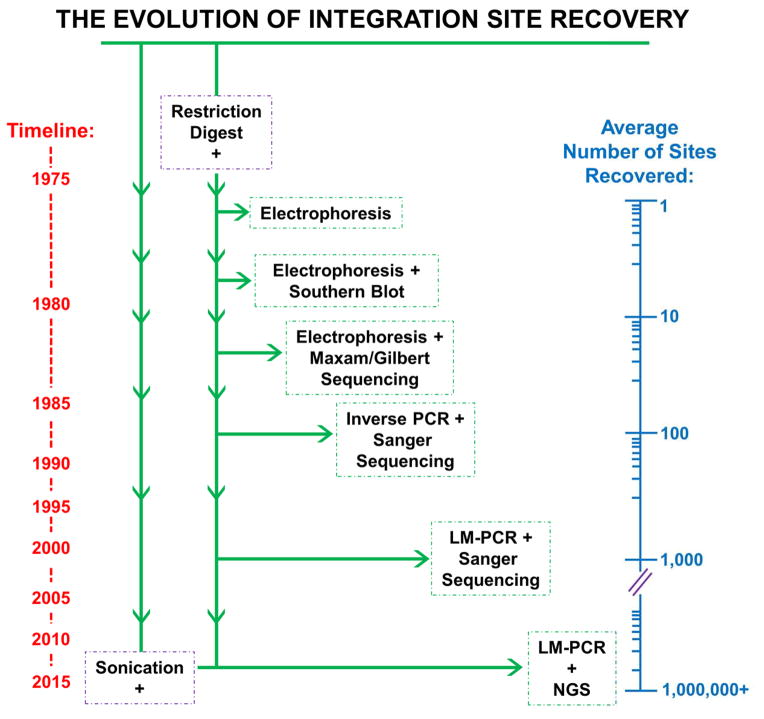
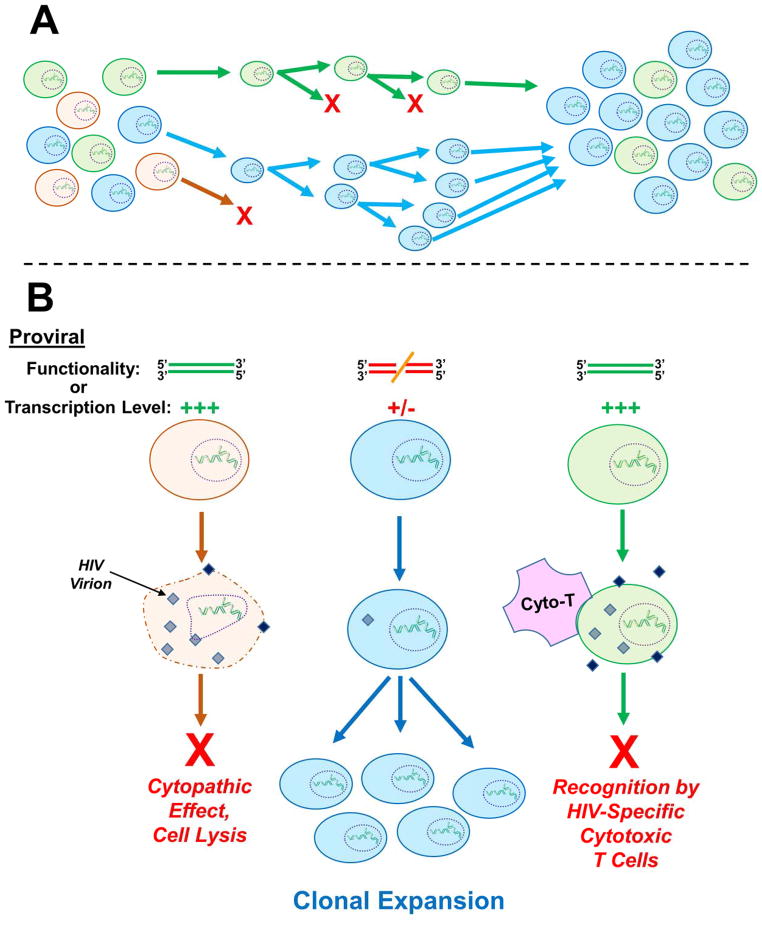
One of the most crucial steps in the life cycle of a retrovirus is the integration of the viral DNA (vDNA) copy of the RNA genome into the genome of an infected host cell. Integration provides for efficient viral gene expression as well as for the segregation of viral genomes to daughter cells upon cell division. Some integrated viruses are not well expressed, and cells latently infected with human immunodeficiency virus type 1 (HIV-1) can resist the action of potent antiretroviral drugs and remain dormant for decades. Intensive research has been dedicated to understanding the catalytic mechanism of integration, as well as the viral and cellular determinants that influence integration site distribution throughout the host genome. In this review, we summarize the evolution of techniques that have been used to recover and map retroviral integration sites, from the early days that first indicated that integration could occur in multiple cellular DNA locations, to current technologies that map upwards of millions of unique integration sites from single in vitro integration reactions or cell culture infections. We further review important insights gained from the use of such mapping techniques, including the monitoring of cell clonal expansion in patients treated with retrovirus-based gene therapy vectors, or patients with acquired immune deficiency syndrome (AIDS) on suppressive antiretroviral therapy (ART). These insights span from integrase (IN) enzyme sequence preferences within target DNA (tDNA) at the sites of integration, to the roles of host cellular proteins in mediating global integration distribution, to the potential relationship between genomic location of vDNA integration site and retroviral latency.
Keywords: Gene therapy; HIV-1; illumina; intasome; integrase; integration sites; next-generation sequencing; retrovirus.
Conflict of interest statement
The authors declare no conflict of financial interest. This work was supported by US National Institutes of Health grants AI052014 and AI039394 (to A.N.E.) as well as AI007386 (to E.S.).
Figures

References
-
- AIUTI A, BIASCO L, SCARAMUZZA S, FERRUA F, CICALESE MP, BARICORDI C, DIONISIO F, CALABRIA A, GIANNELLI S, CASTIELLO MC, BOSTICARDO M, EVANGELIO C, ASSANELLI A, CASIRAGHI M, DI NUNZIO S, CALLEGARO L, BENATI C, RIZZARDI P, PELLIN D, DI SERIO C, SCHMIDT M, VON KALLE C, GARDNER J, MEHTA N, NEDUVA V, DOW DJ, GALY A, MINIERO R, FINOCCHI A, METIN A, BANERJEE PP, ORANGE JS, GALIMBERTI S, VALSECCHI MG, BIFFI A, MONTINI E, VILLA A, CICERI F, RONCAROLO MG, NALDINI L. Lentiviral hematopoietic stem cell gene therapy in patients with Wiskott-Aldrich syndrome. Science. 2013;341:1233151. - PMC - PubMed
-
- AIYER S, SWAPNA GVT, MALANI N, ARAMINI JM, SCHNEIDER WM, PLUMB MR, GHANEM M, LARUE RC, SHARMA A, STUDAMIRE B, KVARATSKHELIA M, BUSHMAN FD, MONTELIONE GT, ROTH MJ. Altering murine leukemia virus integration through disruption of the integrase and BET protein family interaction. Nucleic Acids Res. 2014;42:5917–5928. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources