

Nitrate-Dependent Control of Shoot K Homeostasis by the Nitrate Transporter1/Peptide Transporter Family Member NPF7.3/NRT1.5 and the Stelar K+ Outward Rectifier SKOR in Arabidopsis
- PMID: 26508776
- PMCID: PMC4677904
- DOI: 10.1104/pp.15.01152
Nitrate-Dependent Control of Shoot K Homeostasis by the Nitrate Transporter1/Peptide Transporter Family Member NPF7.3/NRT1.5 and the Stelar K+ Outward Rectifier SKOR in Arabidopsis
Abstract
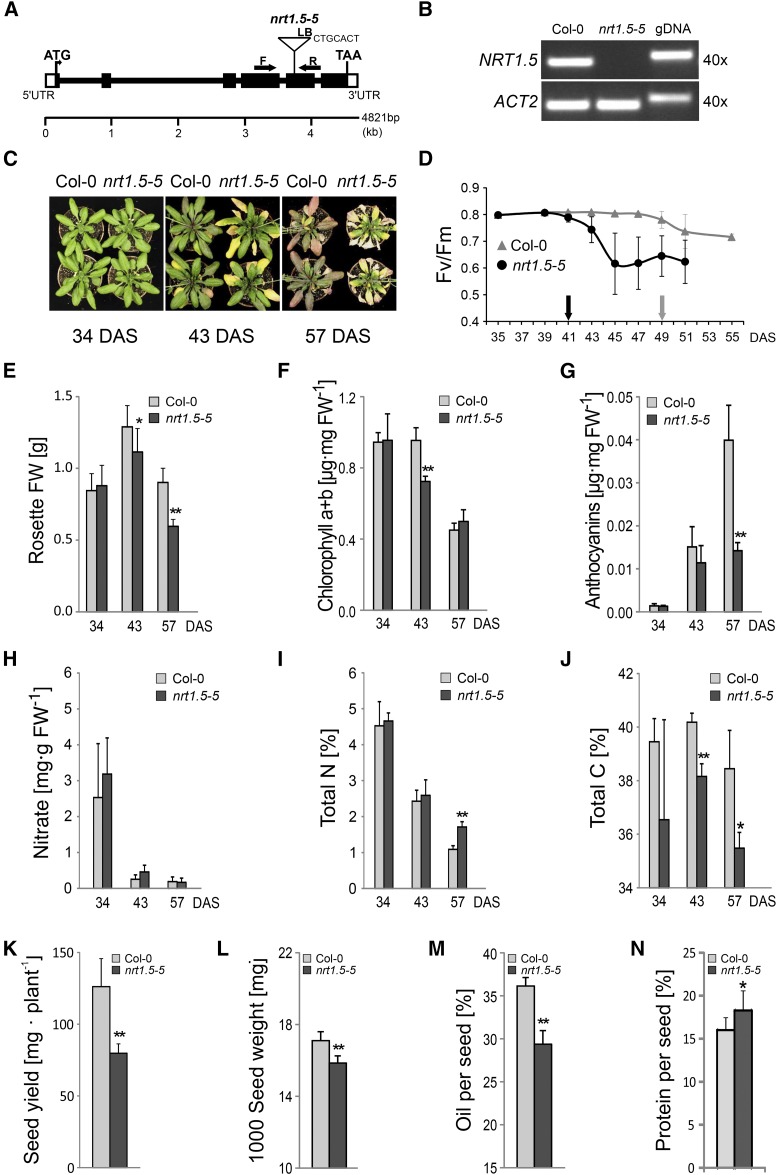
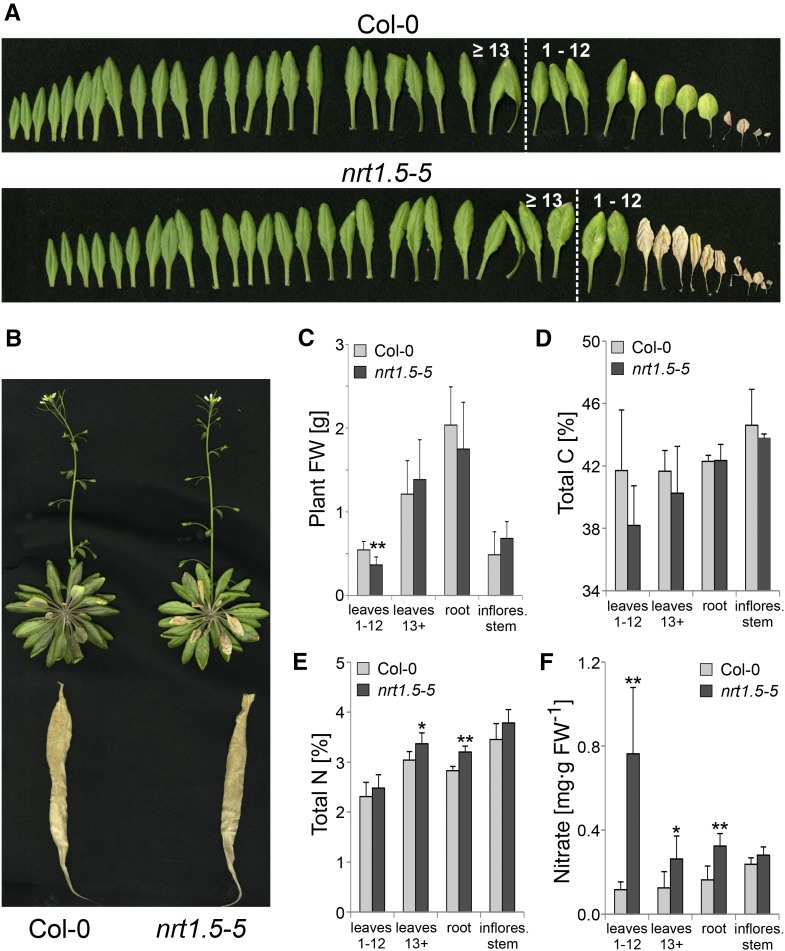
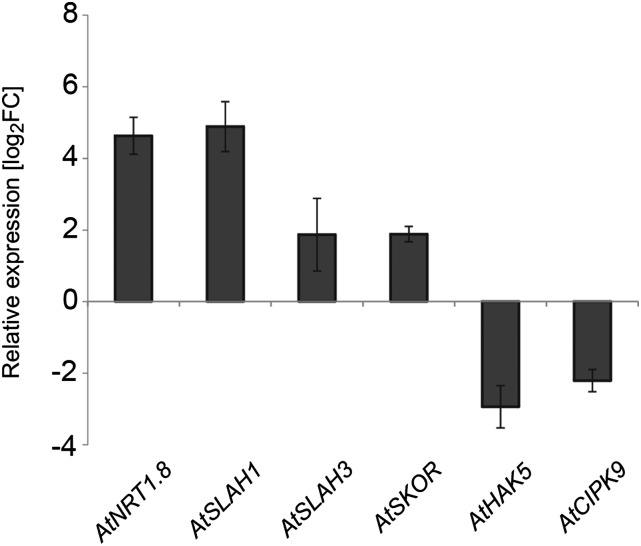
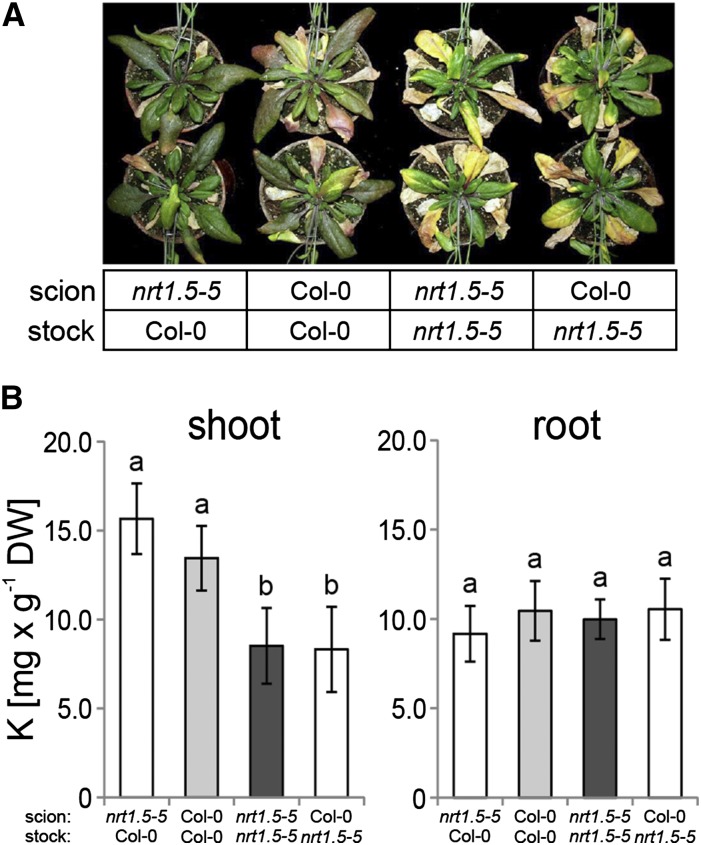
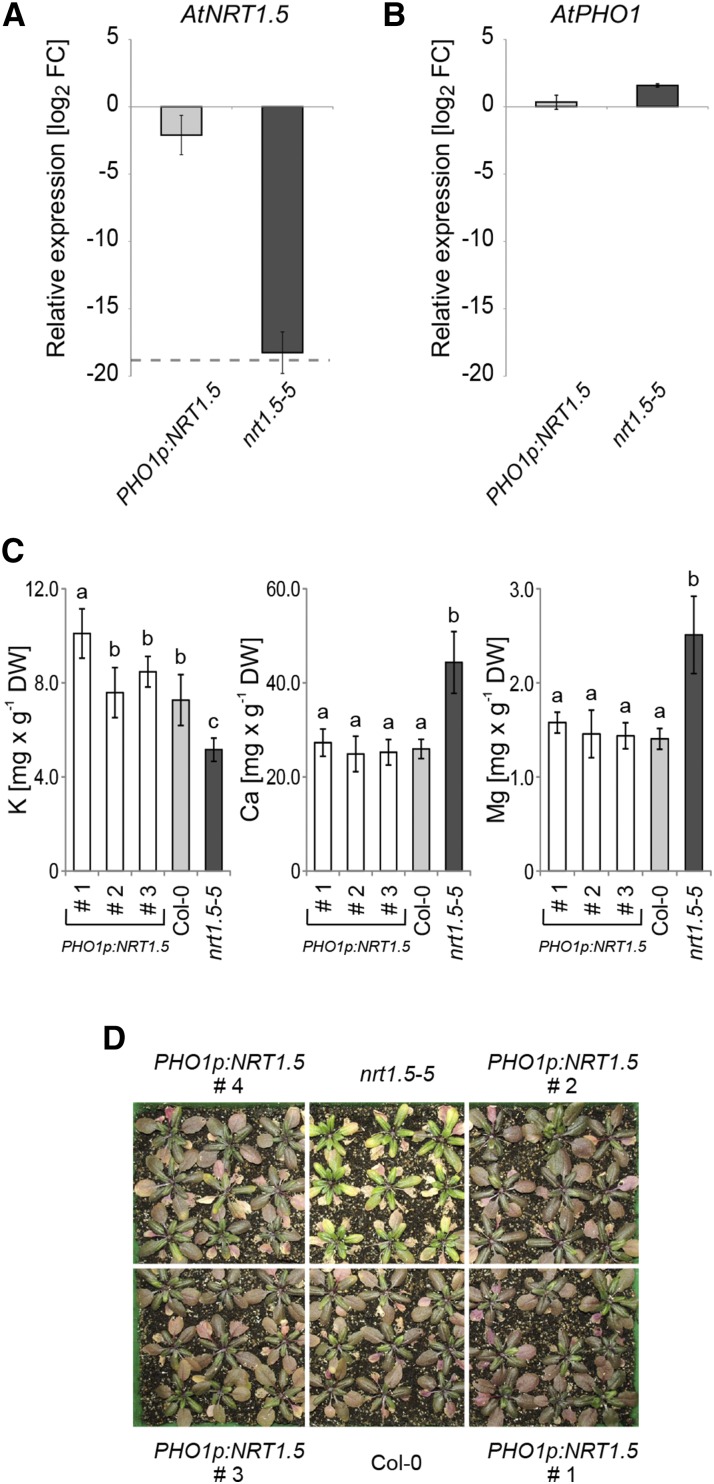
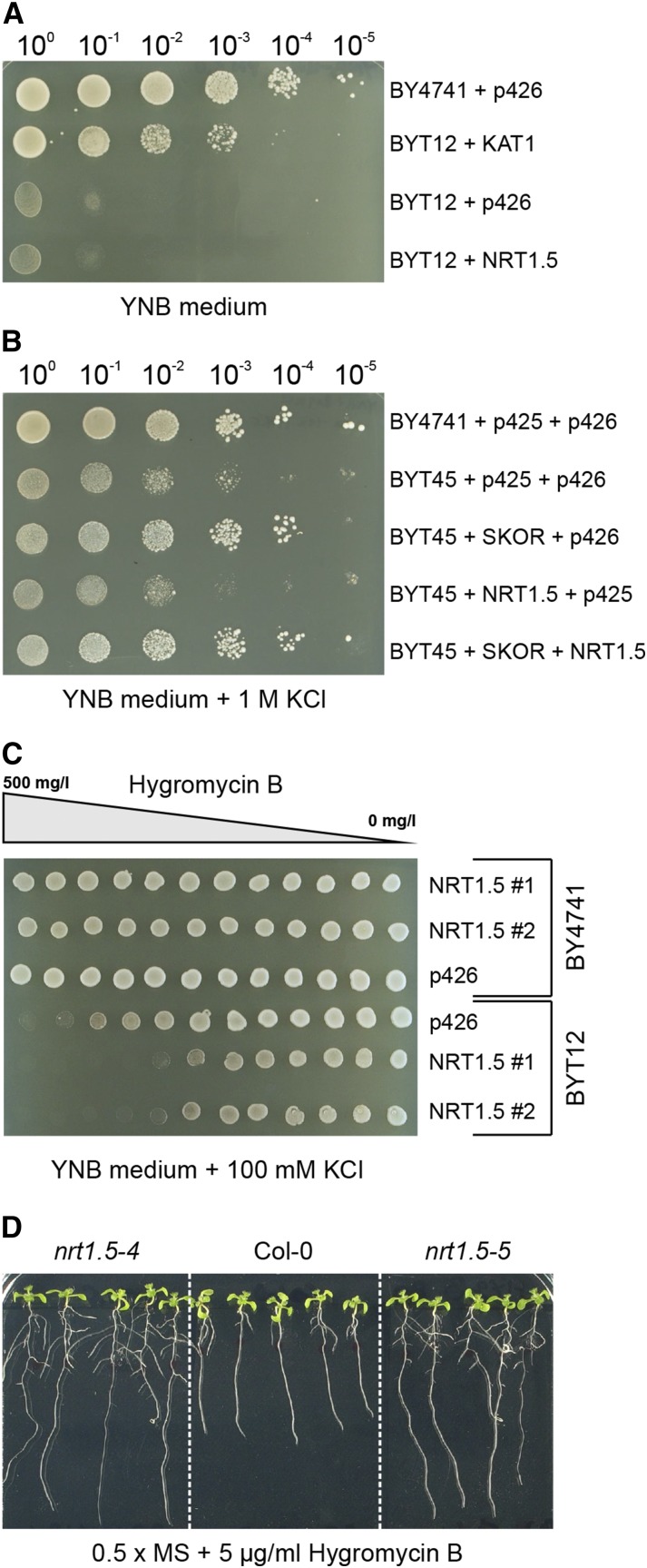
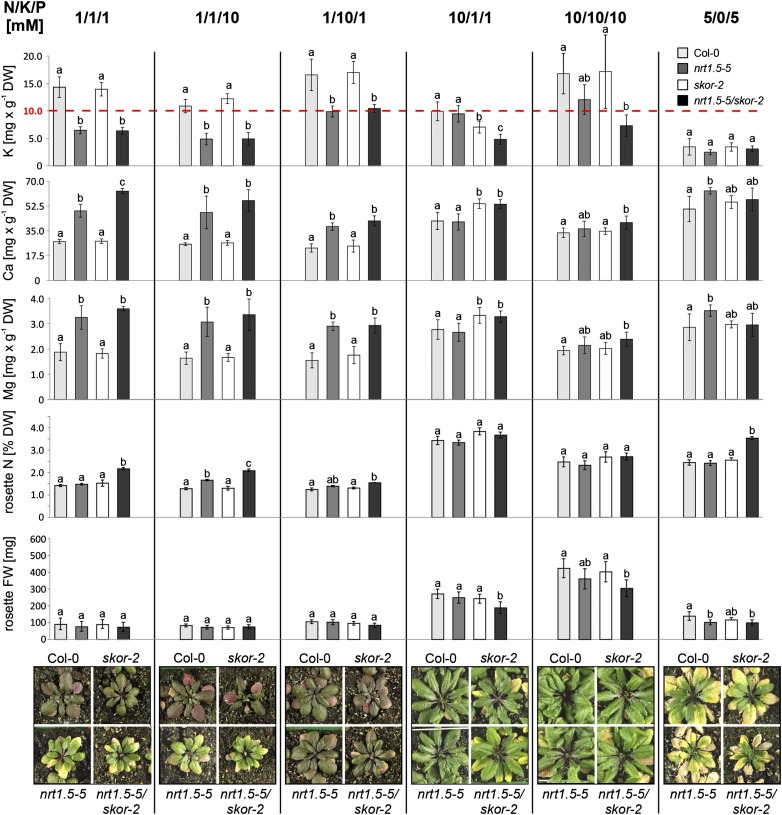
Root-to-shoot translocation and shoot homeostasis of potassium (K) determine nutrient balance, growth, and stress tolerance of vascular plants. To maintain the cation-anion balance, xylem loading of K(+) in the roots relies on the concomitant loading of counteranions, like nitrate (NO3 (-)). However, the coregulation of these loading steps is unclear. Here, we show that the bidirectional, low-affinity Nitrate Transporter1 (NRT1)/Peptide Transporter (PTR) family member NPF7.3/NRT1.5 is important for the NO3 (-)-dependent K(+) translocation in Arabidopsis (Arabidopsis thaliana). Lack of NPF7.3/NRT1.5 resulted in K deficiency in shoots under low NO3 (-) nutrition, whereas the root elemental composition was unchanged. Gene expression data corroborated K deficiency in the nrt1.5-5 shoot, whereas the root responded with a differential expression of genes involved in cation-anion balance. A grafting experiment confirmed that the presence of NPF7.3/NRT1.5 in the root is a prerequisite for proper root-to-shoot translocation of K(+) under low NO3 (-) supply. Because the depolarization-activated Stelar K(+) Outward Rectifier (SKOR) has previously been described as a major contributor for root-to-shoot translocation of K(+) in Arabidopsis, we addressed the hypothesis that NPF7.3/NRT1.5-mediated NO3 (-) translocation might affect xylem loading and root-to-shoot K(+) translocation through SKOR. Indeed, growth of nrt1.5-5 and skor-2 single and double mutants under different K/NO3 (-) regimes revealed that both proteins contribute to K(+) translocation from root to shoot. SKOR activity dominates under high NO3 (-) and low K(+) supply, whereas NPF7.3/NRT1.5 is required under low NO3 (-) availability. This study unravels nutritional conditions as a critical factor for the joint activity of SKOR and NPF7.3/NRT1.5 for shoot K homeostasis.
© 2015 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Bainbridge K, Bennett T, Turnbull C, Leyser O (2006) Grafting. Methods Mol Biol 323: 39–44 - PubMed
-
- Barbier-Brygoo H, De Angeli A, Filleur S, Frachisse JM, Gambale F, Thomine S, Wege S (2011) Anion channels/transporters in plants: from molecular bases to regulatory networks. Annu Rev Plant Biol 62: 25–51 - PubMed
-
- Barneix AJ, Breteler H (1985) Effect of cations on uptake, translocation and reduction of nitrate in wheat seedlings. New Phytol 99: 367–379
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
