

Dynamic membrane protein topological switching upon changes in phospholipid environment
- PMID: 26512118
- PMCID: PMC4653158
- DOI: 10.1073/pnas.1512994112
Dynamic membrane protein topological switching upon changes in phospholipid environment
Abstract
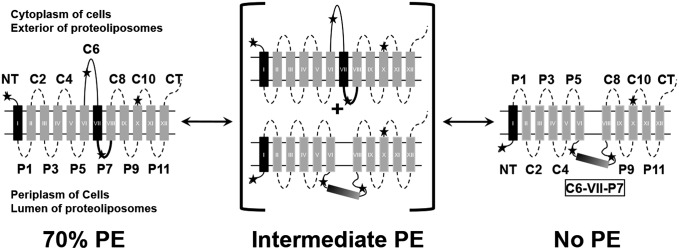
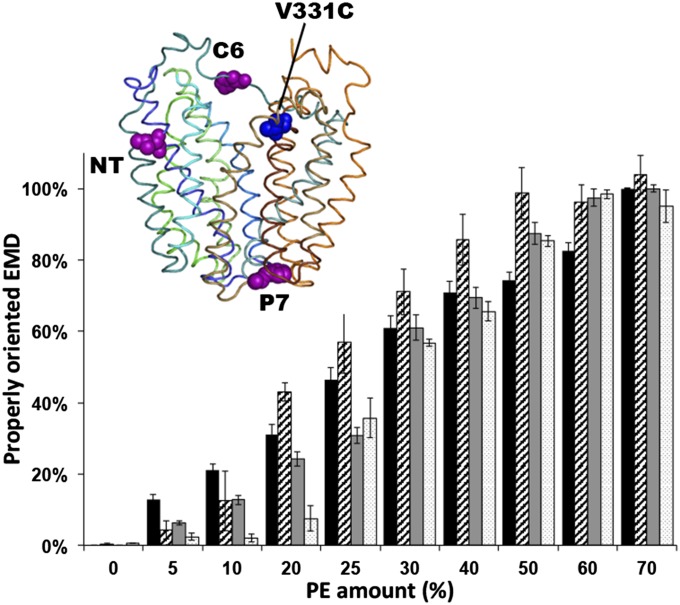
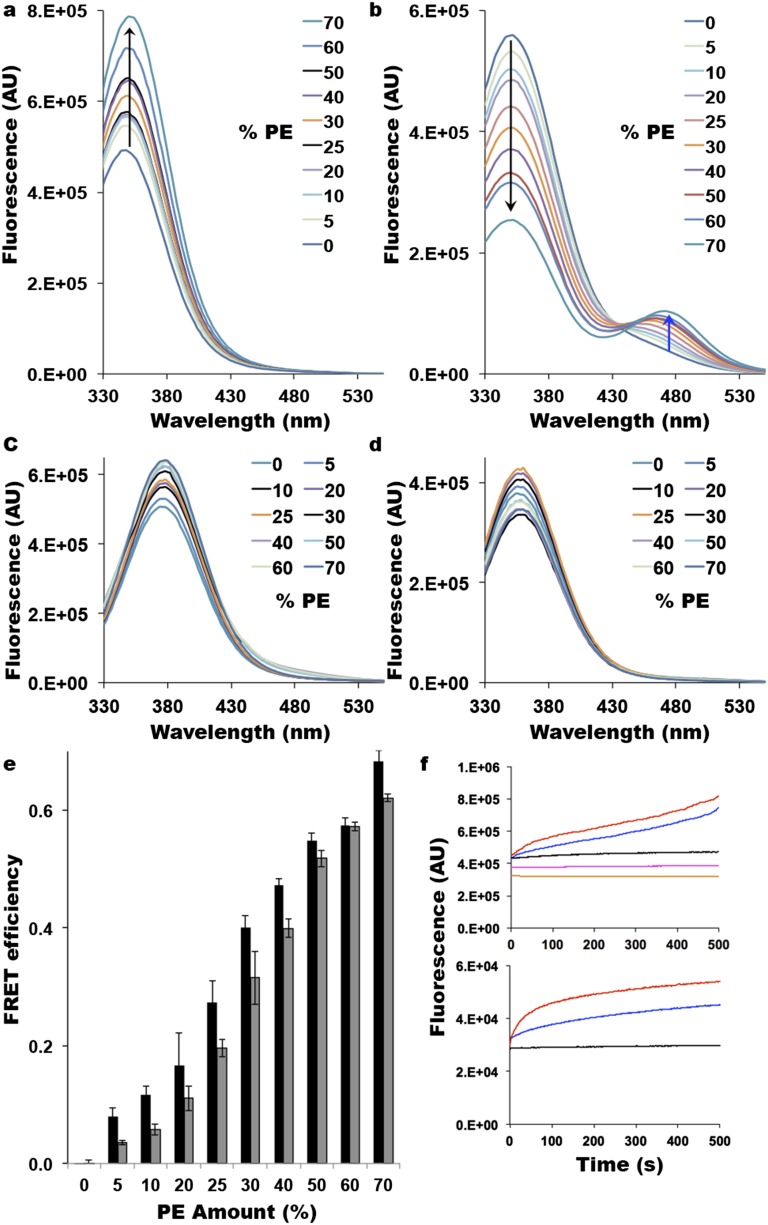
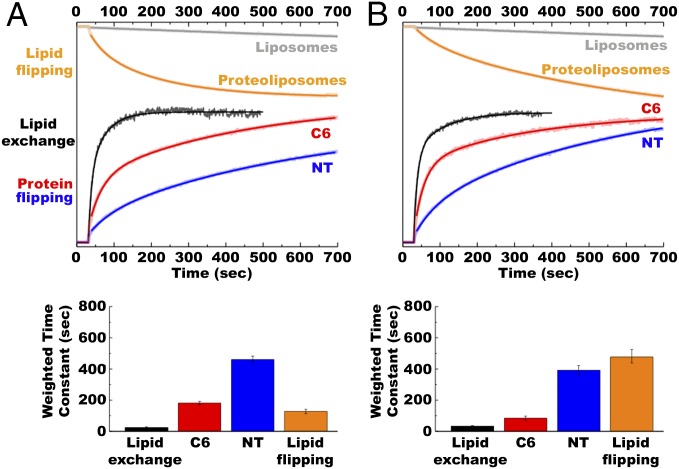
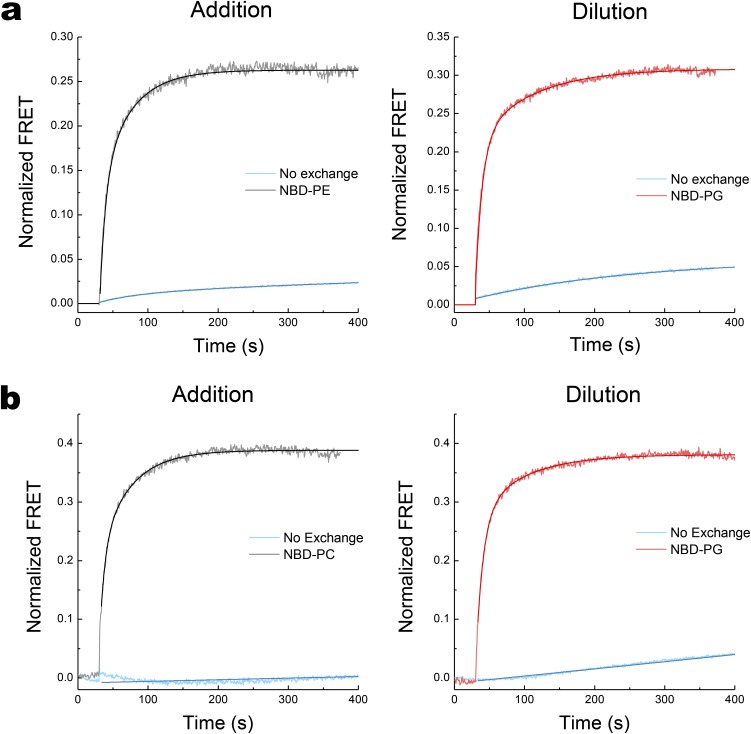
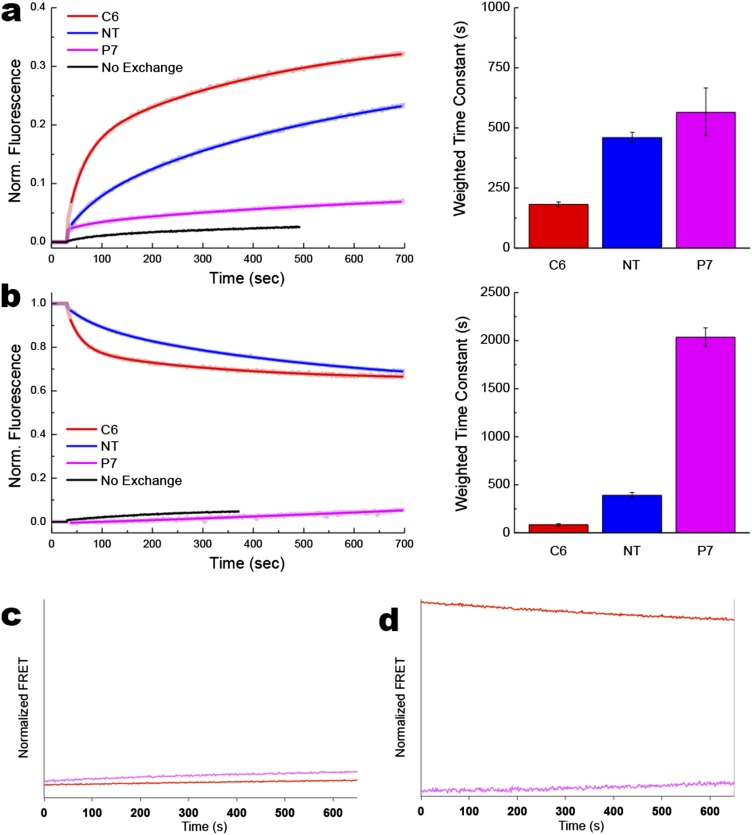
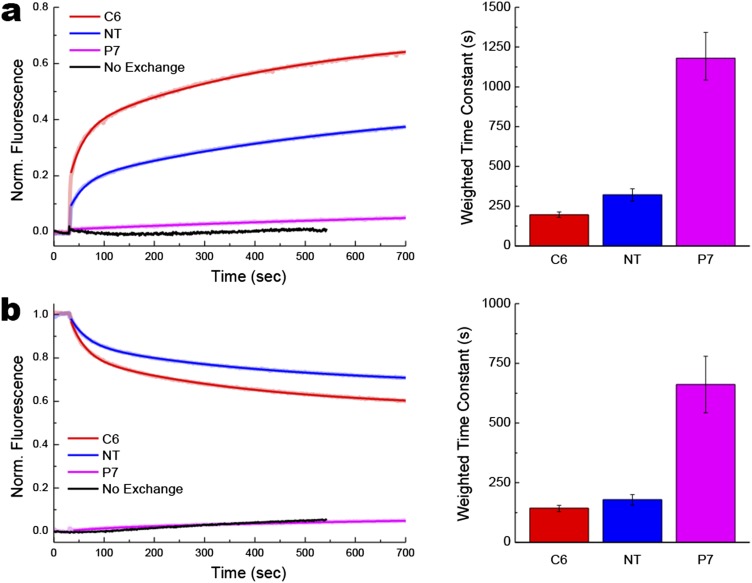
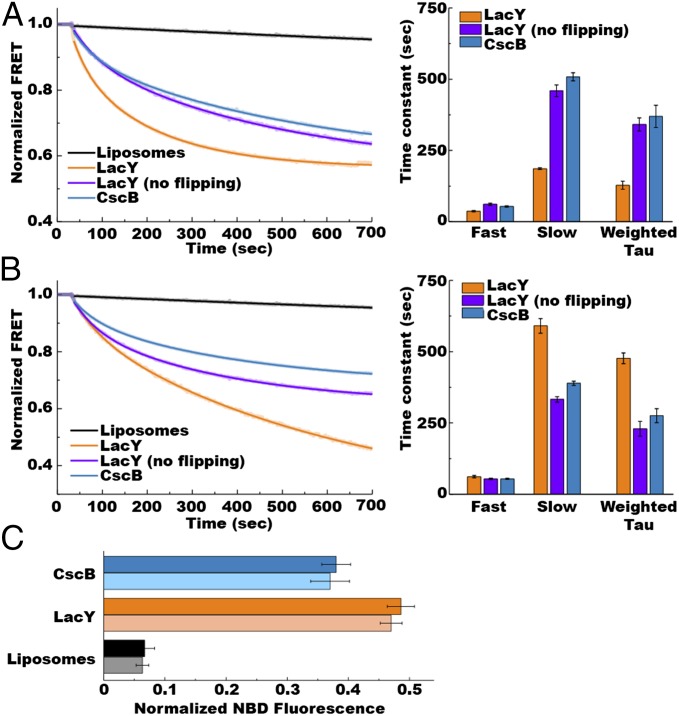
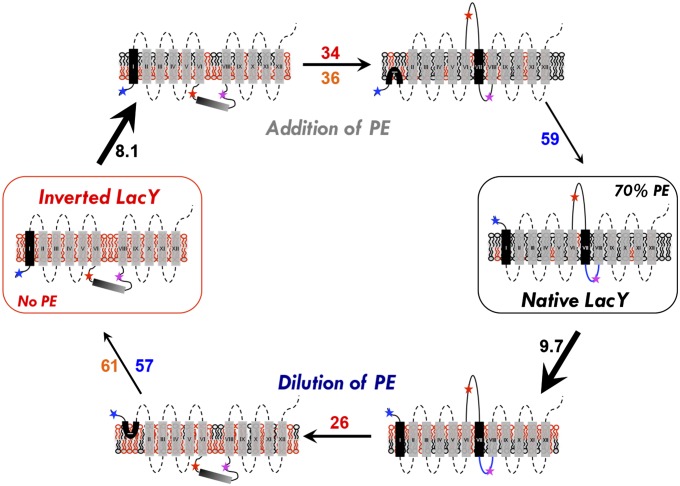
A fundamental objective in membrane biology is to understand and predict how a protein sequence folds and orients in a lipid bilayer. Establishing the principles governing membrane protein folding is central to understanding the molecular basis for membrane proteins that display multiple topologies, the intrinsic dynamic organization of membrane proteins, and membrane protein conformational disorders resulting in disease. We previously established that lactose permease of Escherichia coli displays a mixture of topological conformations and undergoes postassembly bidirectional changes in orientation within the lipid bilayer triggered by a change in membrane phosphatidylethanolamine content, both in vivo and in vitro. However, the physiological implications and mechanism of dynamic structural reorganization of membrane proteins due to changes in lipid environment are limited by the lack of approaches addressing the kinetic parameters of transmembrane protein flipping. In this study, real-time fluorescence spectroscopy was used to determine the rates of protein flipping in the lipid bilayer in both directions and transbilayer flipping of lipids triggered by a change in proteoliposome lipid composition. Our results provide, for the first time to our knowledge, a dynamic picture of these events and demonstrate that membrane protein topological rearrangements in response to lipid modulations occur rapidly following a threshold change in proteoliposome lipid composition. Protein flipping was not accompanied by extensive lipid-dependent unfolding of transmembrane domains. Establishment of lipid bilayer asymmetry was not required but may accelerate the rate of protein flipping. Membrane protein flipping was found to accelerate the rate of transbilayer flipping of lipids.
Keywords: lipid–protein interactions; membrane protein; phospholipids; real-time FRET; topology.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
