

When Physics Takes Over: BAR Proteins and Membrane Curvature
- PMID: 26519988
- PMCID: PMC4831700
- DOI: 10.1016/j.tcb.2015.09.005
When Physics Takes Over: BAR Proteins and Membrane Curvature
Abstract
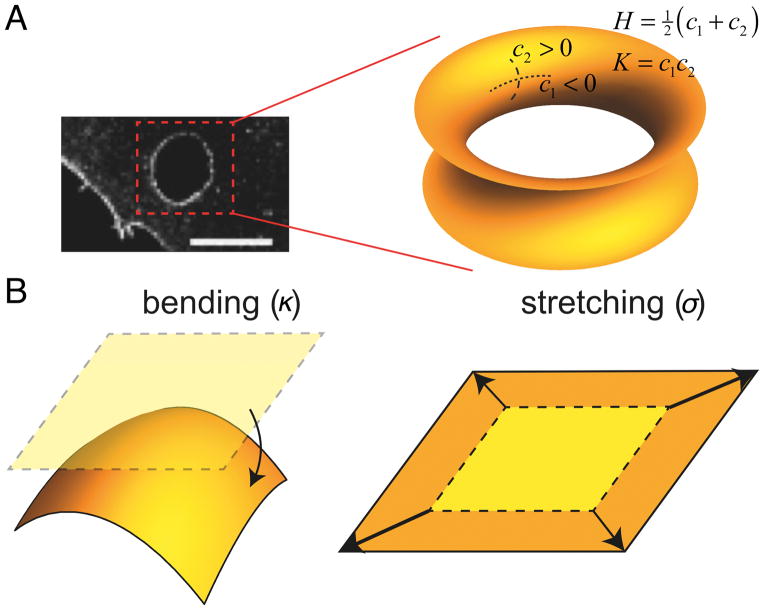
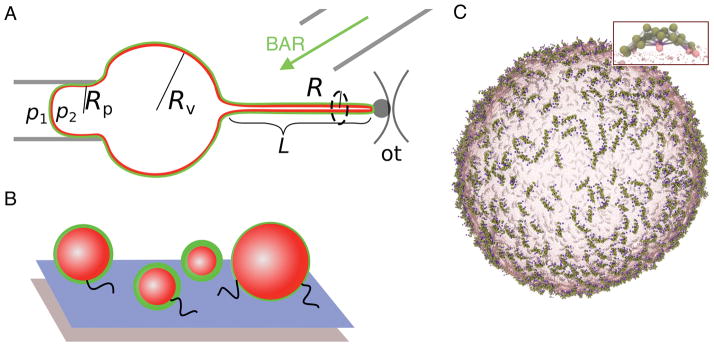
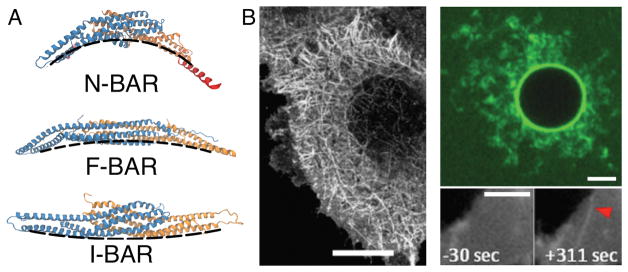
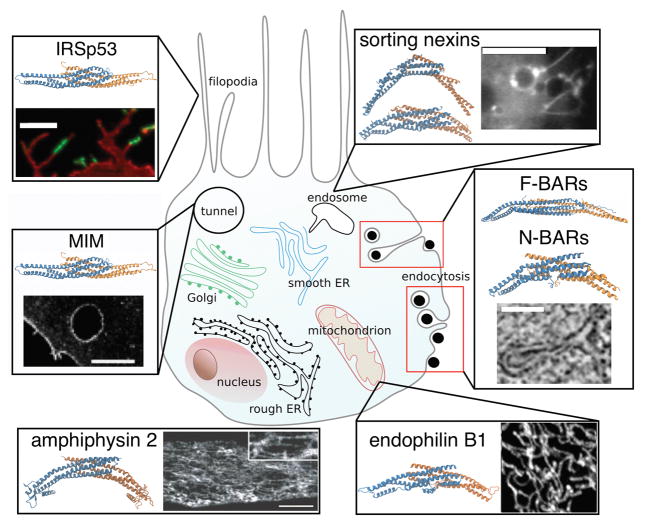
Cell membranes become highly curved during membrane trafficking, cytokinesis, infection, immune response, or cell motion. Bin/amphiphysin/Rvs (BAR) domain proteins with their intrinsically curved and anisotropic shape are involved in many of these processes, but with a large spectrum of modes of action. In vitro experiments and multiscale computer simulations have contributed in identifying a minimal set of physical parameters, namely protein density on the membrane, membrane tension, and membrane shape, that control how bound BAR domain proteins behave on the membrane. In this review, we summarize the multifaceted coupling of BAR proteins to membrane mechanics and propose a simple phase diagram that recapitulates the effects of these parameters.
Keywords: BAR proteins; curvature sensing; membrane curvature; membrane scission; membrane shaping; protein scaffold.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
References
-
- McMahon HT, Gallop JL. Membrane curvature and mechanisms of dynamic cell membrane remodelling. Nature. 2005;438:590–596. - PubMed
-
- Atkins P, de Paula J. Atkins’ Physical Chemistry. OUP; Oxford: 2010.
-
- Goetz R, et al. Mobility and elasticity of self-assembled membranes. Phys Rev Lett. 1999;82:221–224.
-
- Peter BJ, et al. BAR domains as sensors of membrane curvature: the amphiphysin BAR structure. Science. 2004;303:495–499. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
