

Transmembrane protein sorting driven by membrane curvature
- PMID: 26522943
- PMCID: PMC4632190
- DOI: 10.1038/ncomms9728
Transmembrane protein sorting driven by membrane curvature
Abstract
The intricate structure of prokaryotic and eukaryotic cells depends on the ability to target proteins to specific cellular locations. In most cases, we have a poor understanding of the underlying mechanisms. A typical example is the assembly of bacterial chemoreceptors at cell poles. Here we show that the classical chemoreceptor TlpA of Bacillus subtilis does not localize according to the consensus stochastic nucleation mechanism but accumulates at strongly curved membrane areas generated during cell division. This preference was confirmed by accumulation at non-septal curved membranes. Localization appears to be an intrinsic property of the protein complex and does not rely on chemoreceptor clustering, as was previously shown for Escherichia coli. By constructing specific amino-acid substitutions, we demonstrate that the preference for strongly curved membranes arises from the curved shape of chemoreceptor trimer of dimers. These findings demonstrate that the intrinsic shape of transmembrane proteins can determine their cellular localization.
Figures
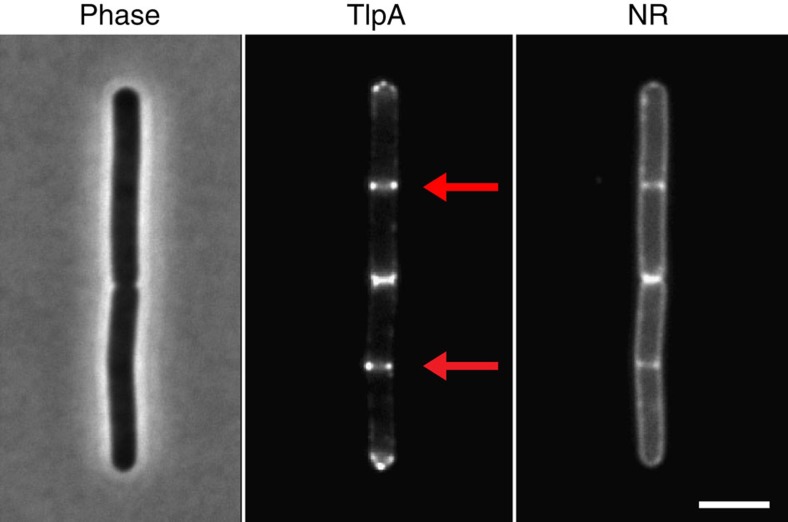
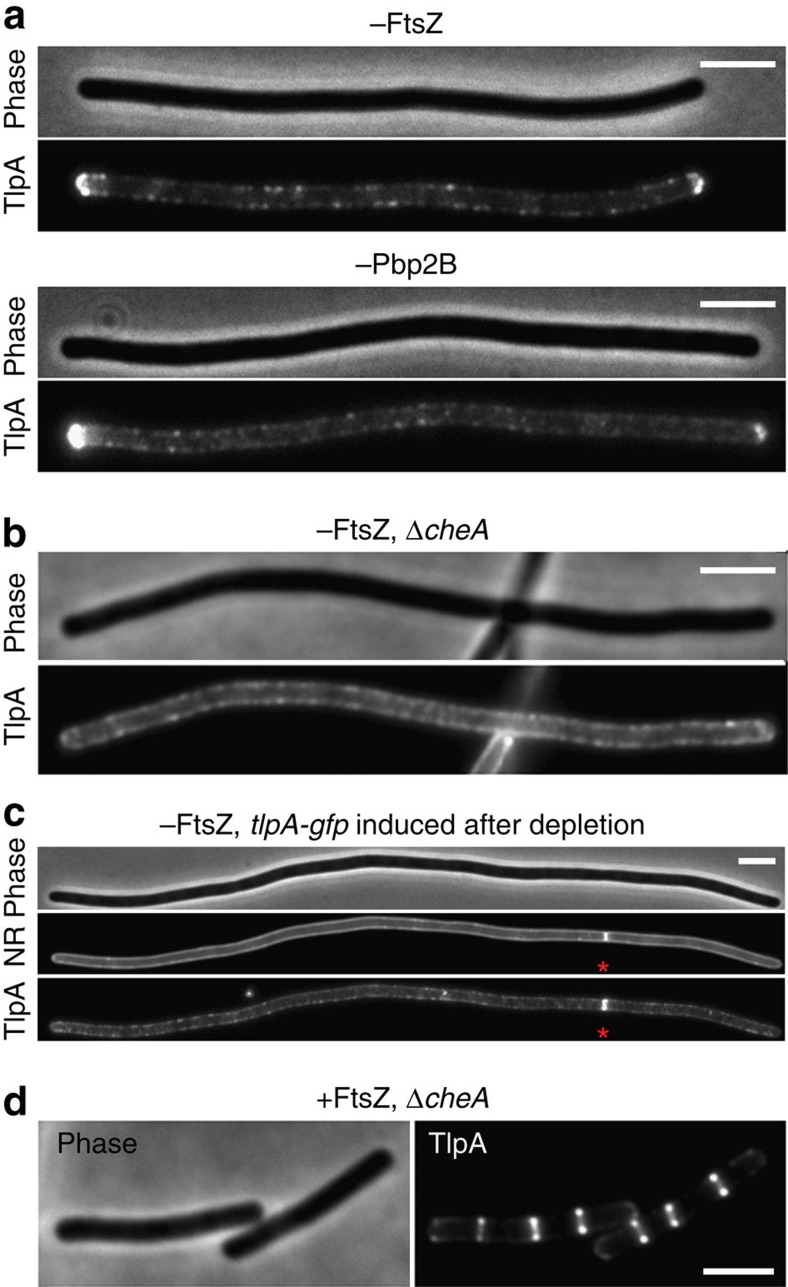
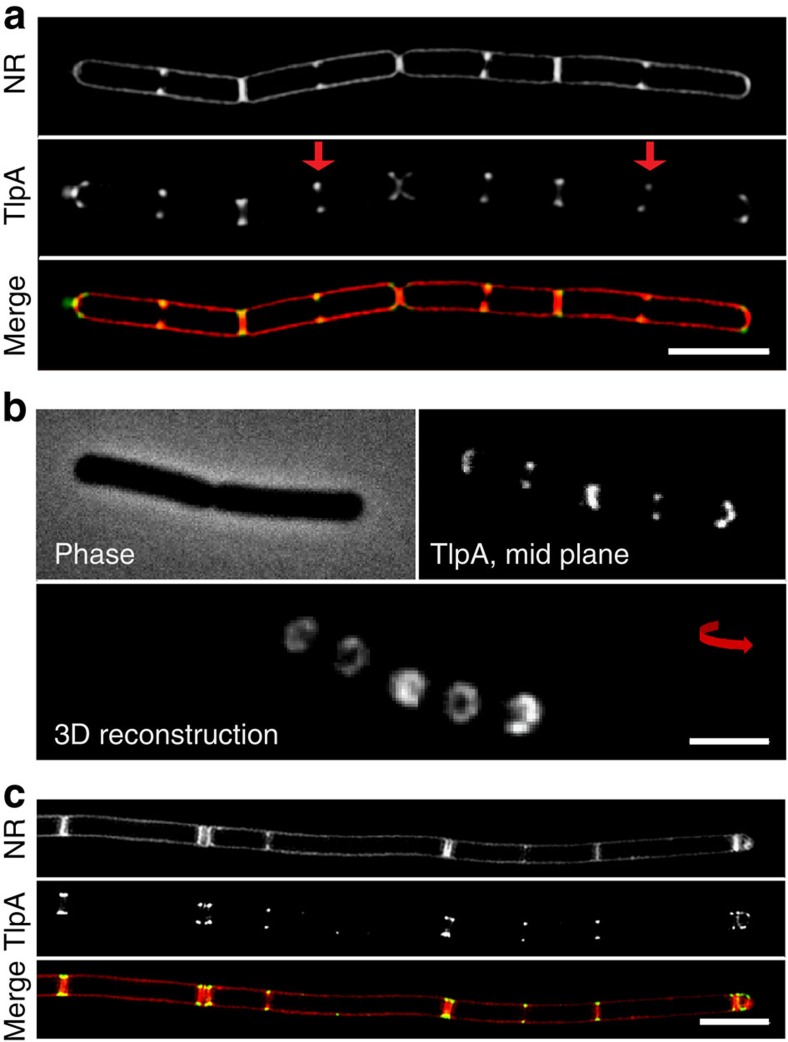
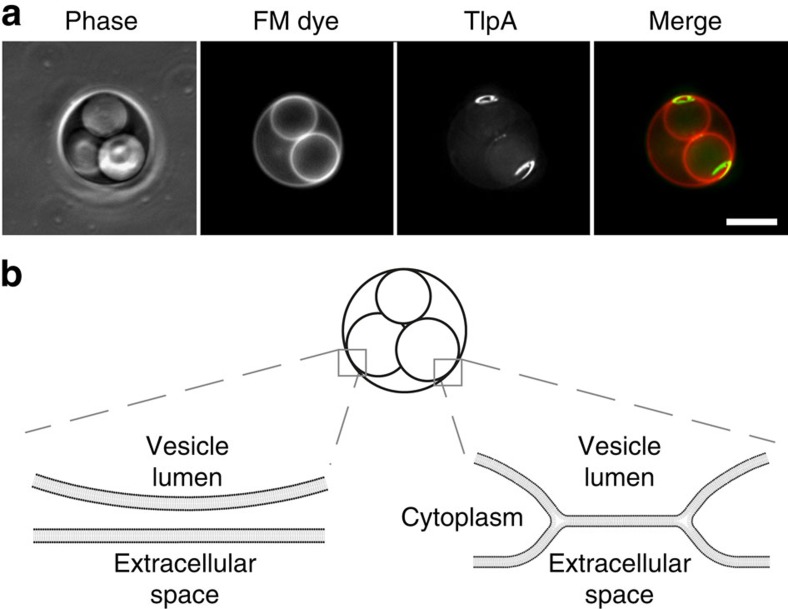
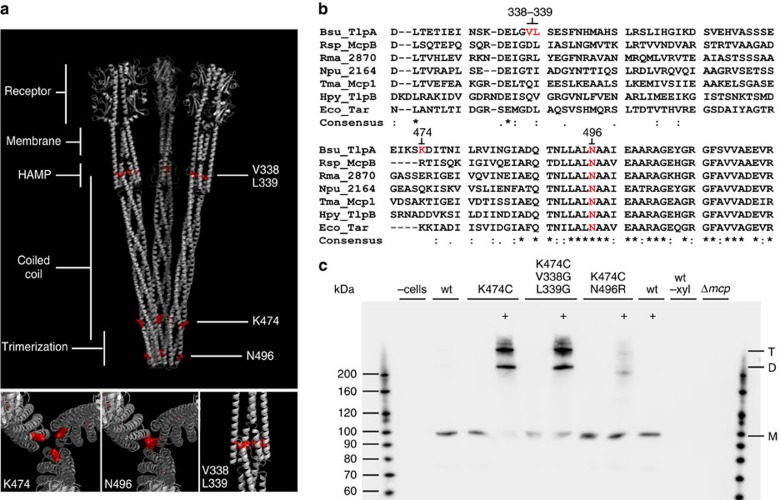
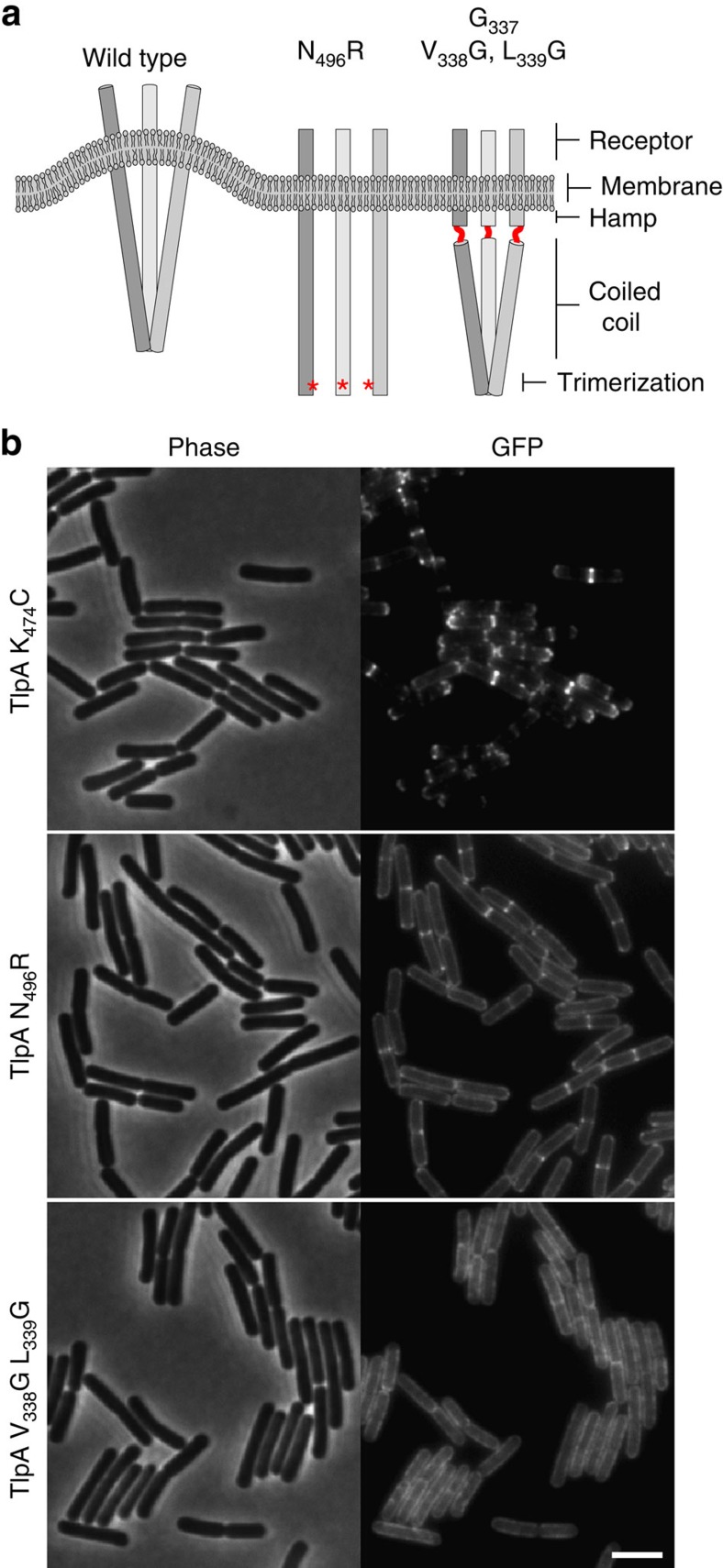
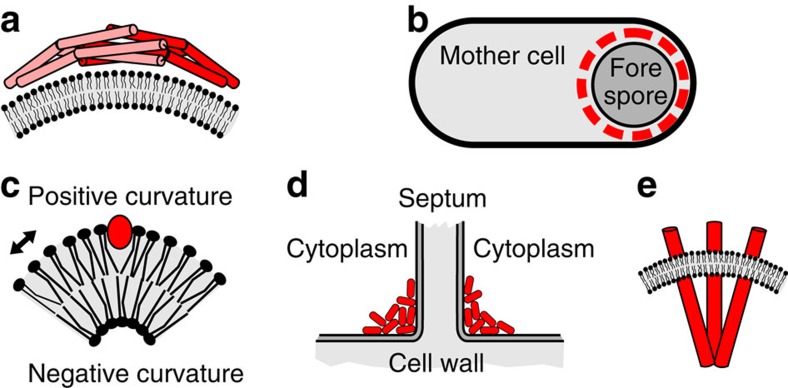
References
-
- Cobbold C., Monaco A. P., Sivaprasadarao A. & Ponnambalam S. Aberrant trafficking of transmembrane proteins in human disease. Trends Cell Biol. 13, 639–647 (2003). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
