

The Corticohippocampal Circuit, Synaptic Plasticity, and Memory
- PMID: 26525152
- PMCID: PMC4632668
- DOI: 10.1101/cshperspect.a021733
The Corticohippocampal Circuit, Synaptic Plasticity, and Memory
Abstract
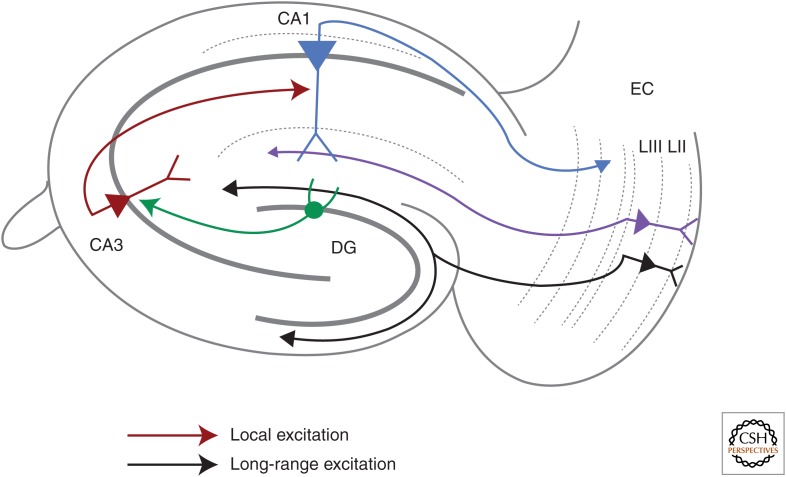
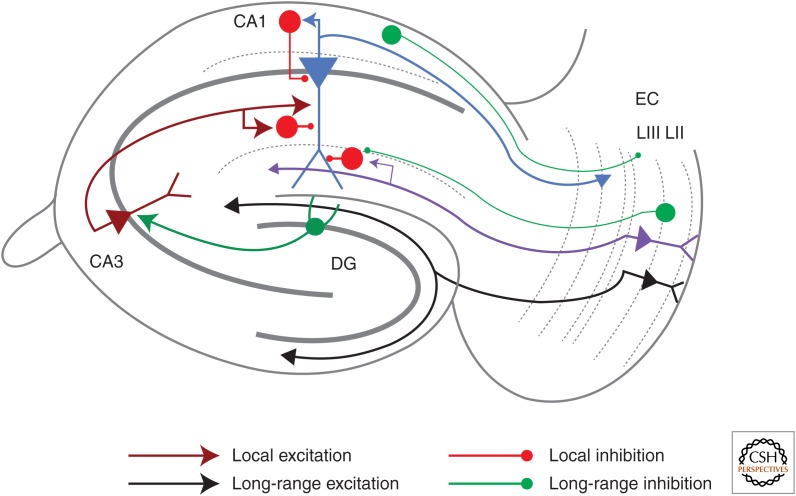
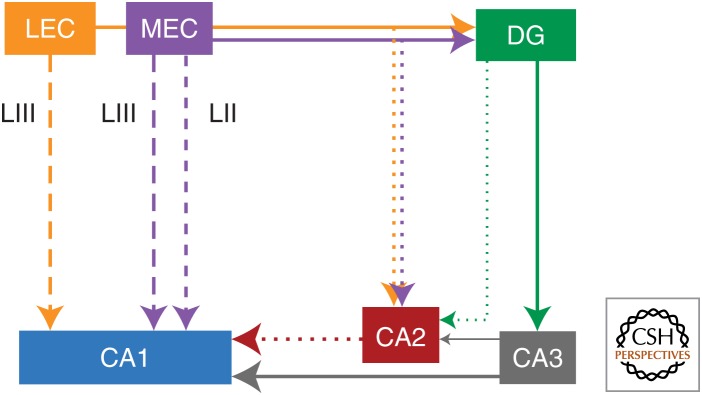
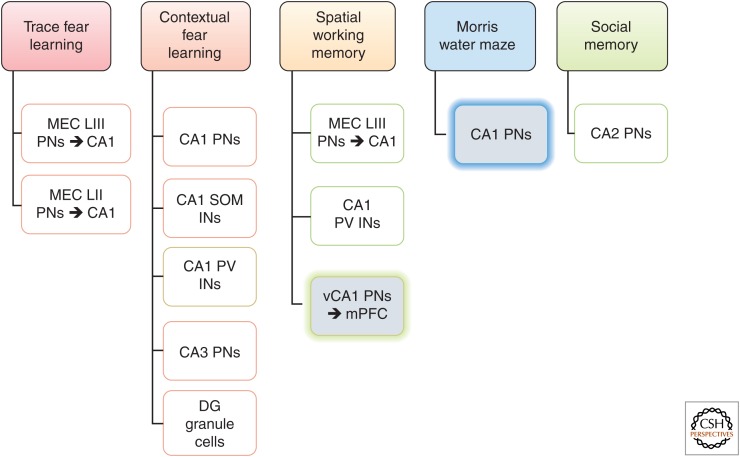
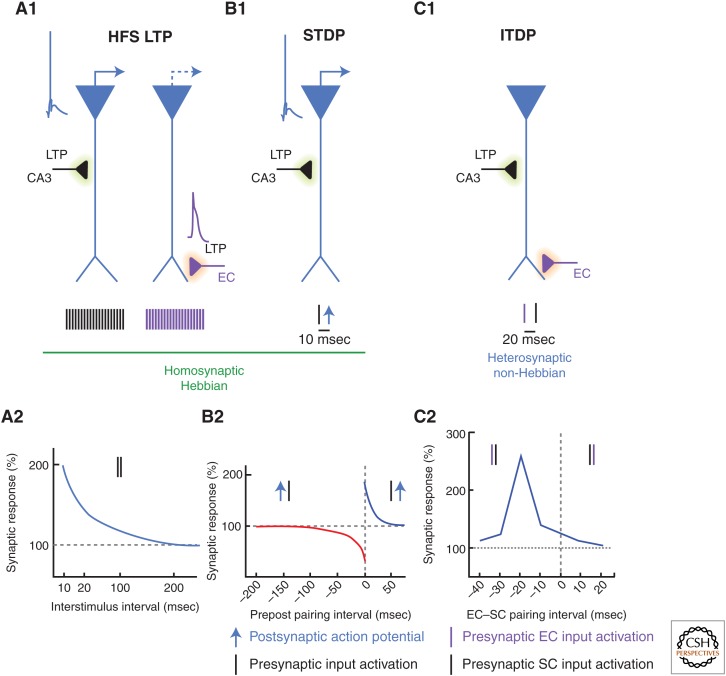
Synaptic plasticity serves as a cellular substrate for information storage in the central nervous system. The entorhinal cortex (EC) and hippocampus are interconnected brain areas supporting basic cognitive functions important for the formation and retrieval of declarative memories. Here, we discuss how information flow in the EC-hippocampal loop is organized through circuit design. We highlight recently identified corticohippocampal and intrahippocampal connections and how these long-range and local microcircuits contribute to learning. This review also describes various forms of activity-dependent mechanisms that change the strength of corticohippocampal synaptic transmission. A key point to emerge from these studies is that patterned activity and interaction of coincident inputs gives rise to associational plasticity and long-term regulation of information flow. Finally, we offer insights about how learning-related synaptic plasticity within the corticohippocampal circuit during sensory experiences may enable adaptive behaviors for encoding spatial, episodic, social, and contextual memories.
Copyright © 2015 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
-
- Abel T, Nguyen PV, Barad M, Deuel TA, Kandel ER, Bourtchouladze R. 1997. Genetic demonstration of a role for PKA in the late phase of LTP and in hippocampus-based long-term memory. Cell 88: 615–626. - PubMed
-
- Abraham WC, Tate WP. 1997. Metaplasticity: A new vista across the field of synaptic plasticity. Prog Neurobiol 52: 303–323. - PubMed
-
- Aihara T, Tsukada M, Crair MC, Shinomoto S. 1997. Stimulus-dependent induction of long-term potentiation in CA1 area of the hippocampus: Experiment and model. Hippocampus 7: 416–426. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical