

CRISPR/dCas9-mediated Transcriptional Inhibition Ameliorates the Epigenetic Dysregulation at D4Z4 and Represses DUX4-fl in FSH Muscular Dystrophy
- PMID: 26527377
- PMCID: PMC4786914
- DOI: 10.1038/mt.2015.200
CRISPR/dCas9-mediated Transcriptional Inhibition Ameliorates the Epigenetic Dysregulation at D4Z4 and Represses DUX4-fl in FSH Muscular Dystrophy
Abstract
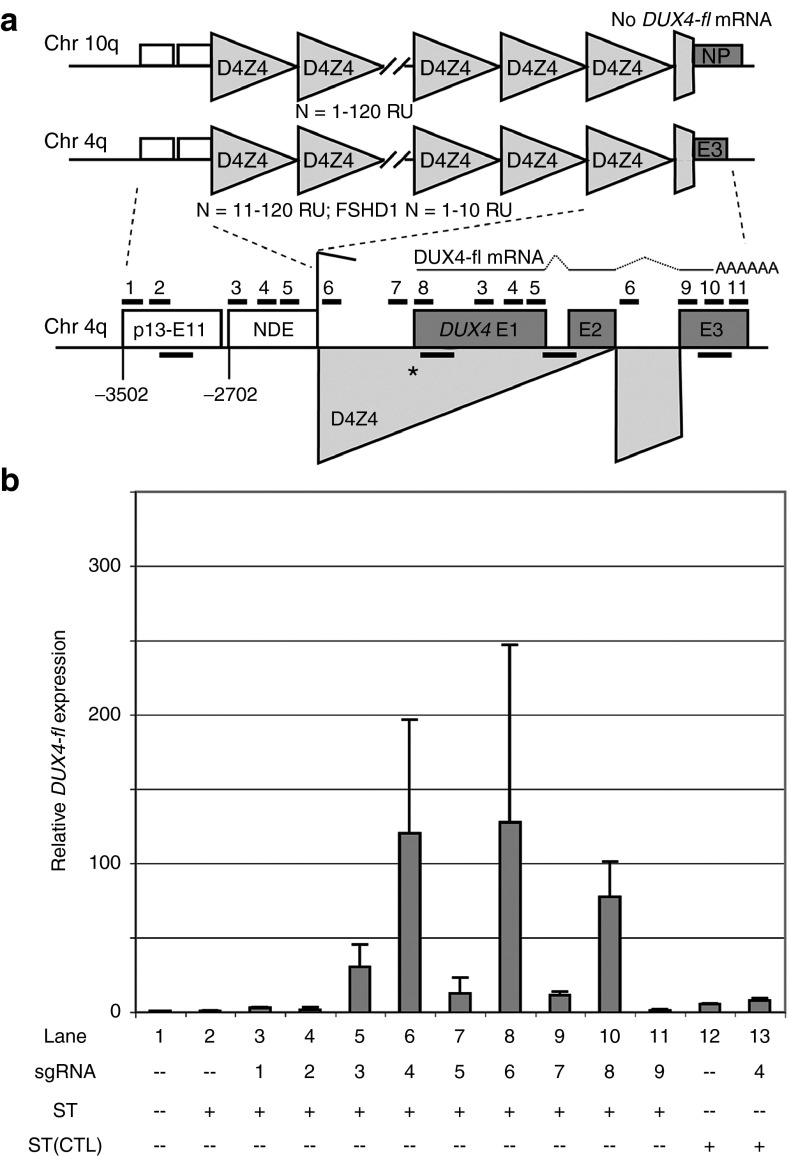
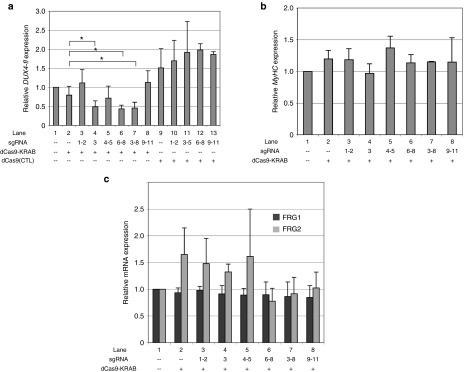
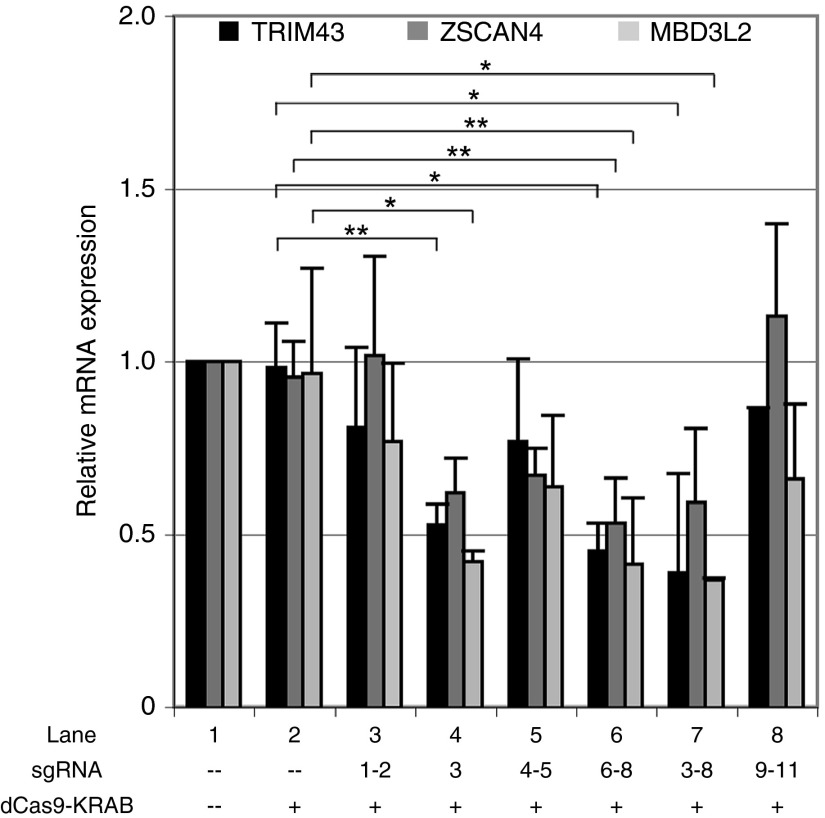
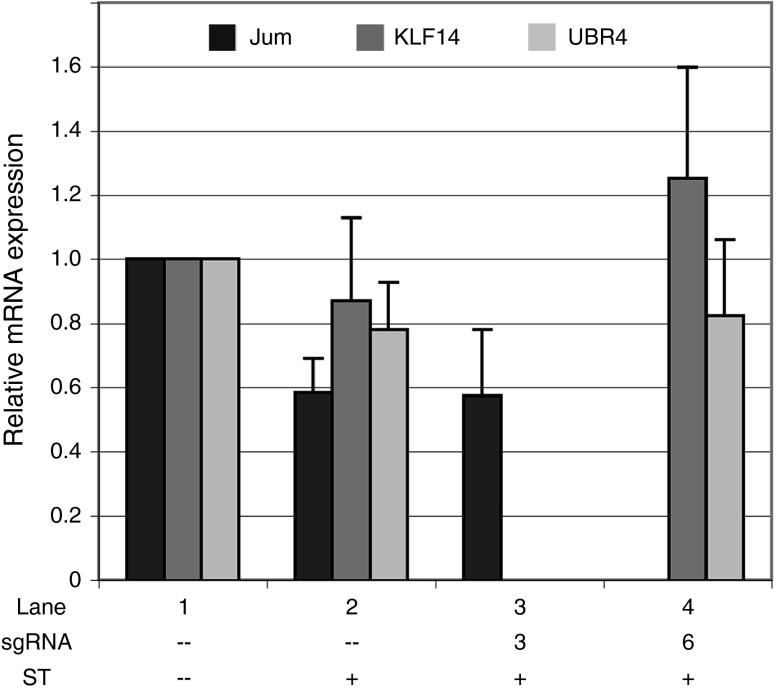
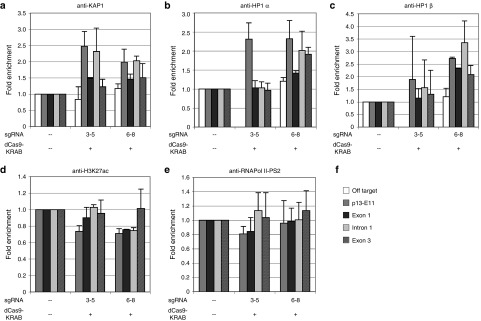
Facioscapulohumeral muscular dystrophy (FSHD) is one of the most prevalent myopathies, affecting males and females of all ages. Both forms of the disease are linked by epigenetic derepression of the D4Z4 macrosatellite repeat array at chromosome 4q35, leading to aberrant expression of D4Z4-encoded RNAs in skeletal muscle. Production of full-length DUX4 (DUX4-fl) mRNA from the derepressed D4Z4 array results in misexpression of DUX4-FL protein and its transcriptional targets, and apoptosis, ultimately leading to accumulated muscle pathology. Returning the chromatin at the FSHD locus to its nonpathogenic, epigenetically repressed state would simultaneously affect all D4Z4 RNAs, inhibiting downstream pathogenic pathways, and is thus an attractive therapeutic strategy. Advances in CRISPR/Cas9-based genome editing make it possible to target epigenetic modifiers to an endogenous disease locus, although reports to date have focused on more typical genomic regions. Here, we demonstrate that a CRISPR/dCas9 transcriptional inhibitor can be specifically targeted to the highly repetitive FSHD macrosatellite array and alter the chromatin to repress expression of DUX4-fl in primary FSHD myocytes. These results implicate the promoter and exon 1 of DUX4 as potential therapeutic targets and demonstrate the utility of CRISPR technology for correction of the epigenetic dysregulation in FSHD.
Figures
References
-
- Orphanet. 2011. Prevalence of rare diseases: bibliographic data in Orphanet report series: rare diseases collection. http://www.orpha.net/orphacom/cahiers/docs/GB/Prevalence_of_rare_disease....
-
- Padberg, GW. Facioscapulohumeral Disease [Thesis]. Leiden University, Leiden, the Netherlands, 1982. pp. 243.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
