

Identification of a novel cis-regulatory element essential for immune tolerance
- PMID: 26527800
- PMCID: PMC4647269
- DOI: 10.1084/jem.20151069
Identification of a novel cis-regulatory element essential for immune tolerance
Abstract
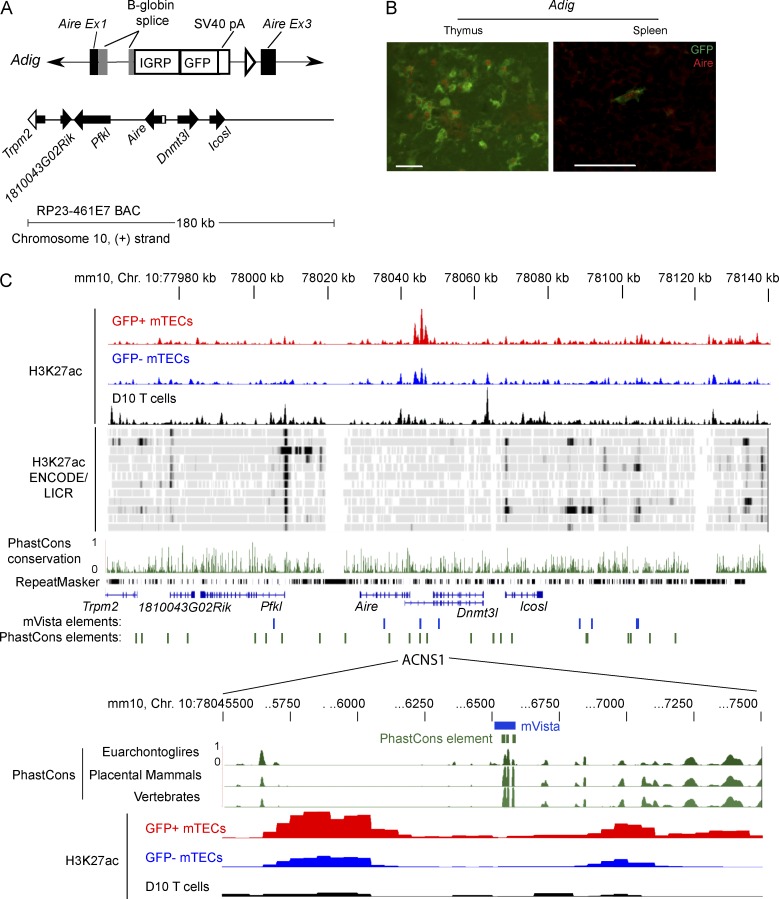
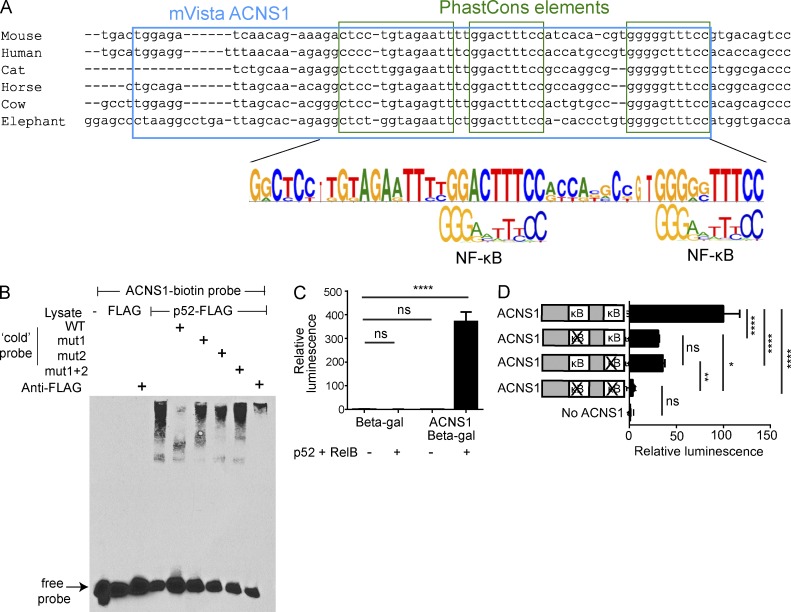
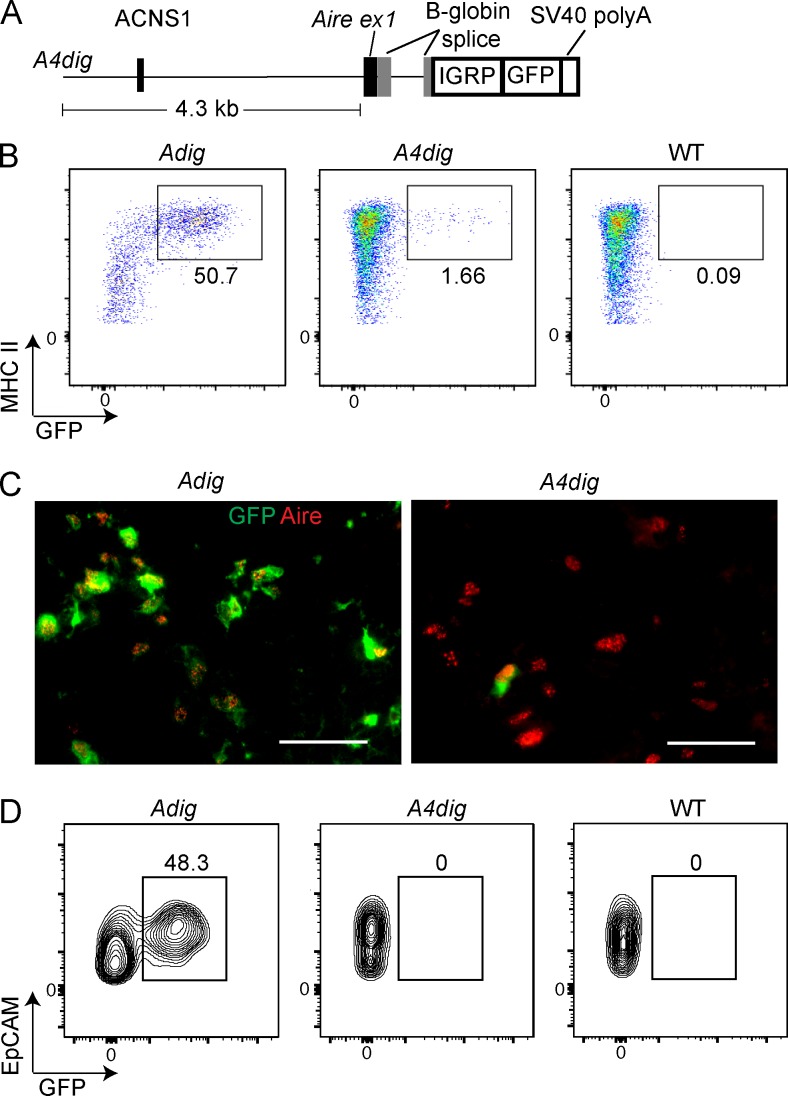
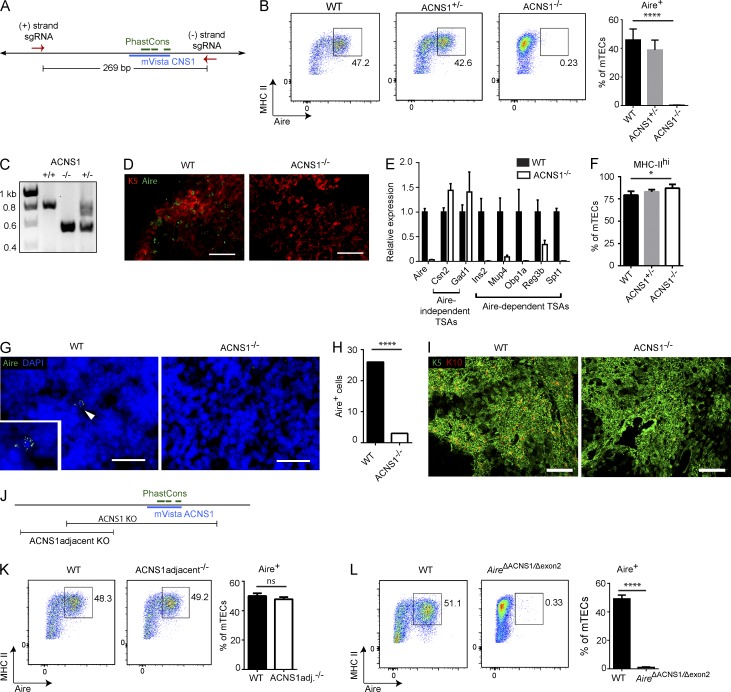
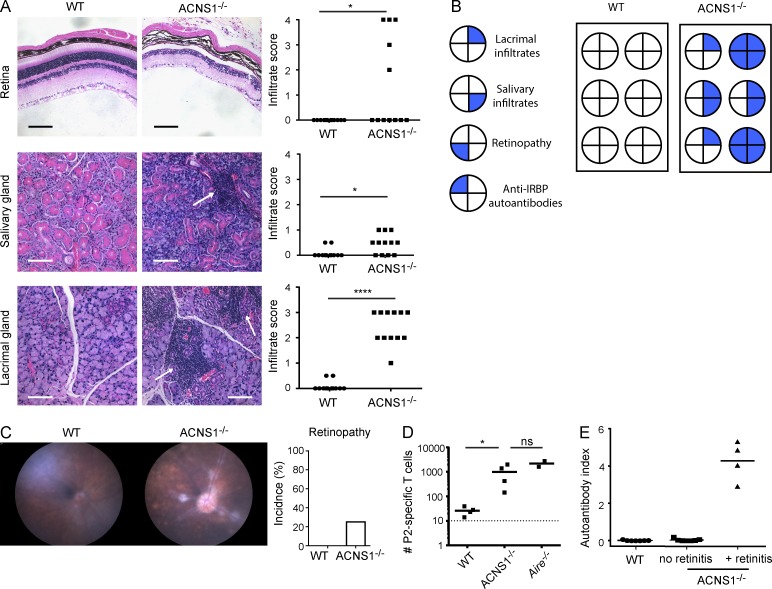
Thymic central tolerance is essential to preventing autoimmunity. In medullary thymic epithelial cells (mTECs), the Autoimmune regulator (Aire) gene plays an essential role in this process by driving the expression of a diverse set of tissue-specific antigens (TSAs), which are presented and help tolerize self-reactive thymocytes. Interestingly, Aire has a highly tissue-restricted pattern of expression, with only mTECs and peripheral extrathymic Aire-expressing cells (eTACs) known to express detectable levels in adults. Despite this high level of tissue specificity, the cis-regulatory elements that control Aire expression have remained obscure. Here, we identify a highly conserved noncoding DNA element that is essential for Aire expression. This element shows enrichment of enhancer-associated histone marks in mTECs and also has characteristics of being an NF-κB-responsive element. Finally, we find that this element is essential for Aire expression in vivo and necessary to prevent spontaneous autoimmunity, reflecting the importance of this regulatory DNA element in promoting immune tolerance.
© 2015 LaFlam et al.
Figures
References
-
- Aaltonen J., Bjorses P., Perheentupa J., Horelli-Kuitunen N., Palotie A., Peltonen L., Lee Y.S., Francis F., Henning S., Thiel C., et al. Finnish-German APECED Consortium . 1997. An autoimmune disease, APECED, caused by mutations in a novel gene featuring two PHD-type zinc-finger domains. Nat. Genet. 17:399–403. 10.1038/ng1297-399 - DOI - PubMed
-
- Akiyama T., Shimo Y., Yanai H., Qin J., Ohshima D., Maruyama Y., Asaumi Y., Kitazawa J., Takayanagi H., Penninger J.M., et al. 2008. The tumor necrosis factor family receptors RANK and CD40 cooperatively establish the thymic medullary microenvironment and self-tolerance. Immunity. 29:423–437. 10.1016/j.immuni.2008.06.015 - DOI - PubMed
-
- Creyghton M.P., Cheng A.W., Welstead G.G., Kooistra T., Carey B.W., Steine E.J., Hanna J., Lodato M.A., Frampton G.M., Sharp P.A., et al. 2010. Histone H3K27ac separates active from poised enhancers and predicts developmental state. Proc. Natl. Acad. Sci. USA. 107:21931–21936. 10.1073/pnas.1016071107 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
