

Loss of CD24 in Mice Leads to Metabolic Dysfunctions and a Reduction in White Adipocyte Tissue
- PMID: 26536476
- PMCID: PMC4633231
- DOI: 10.1371/journal.pone.0141966
Loss of CD24 in Mice Leads to Metabolic Dysfunctions and a Reduction in White Adipocyte Tissue
Abstract
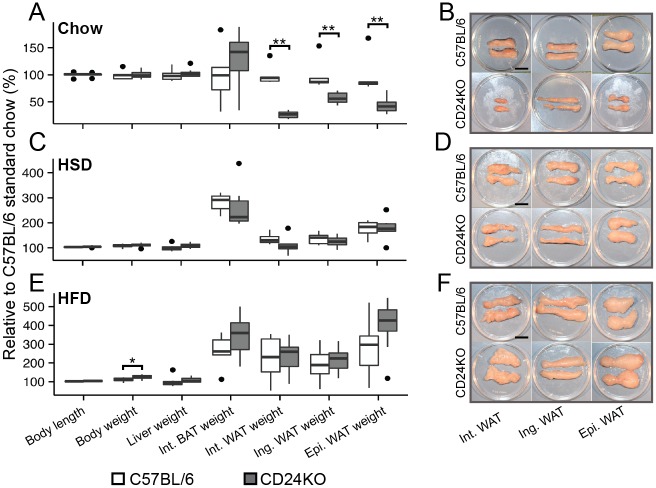
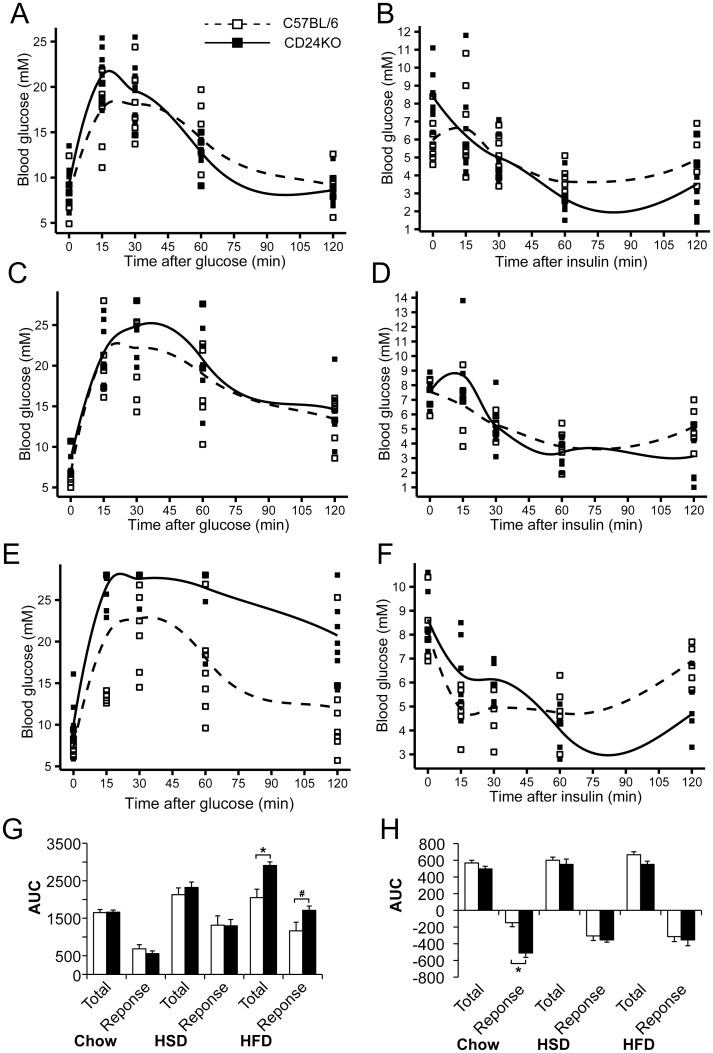
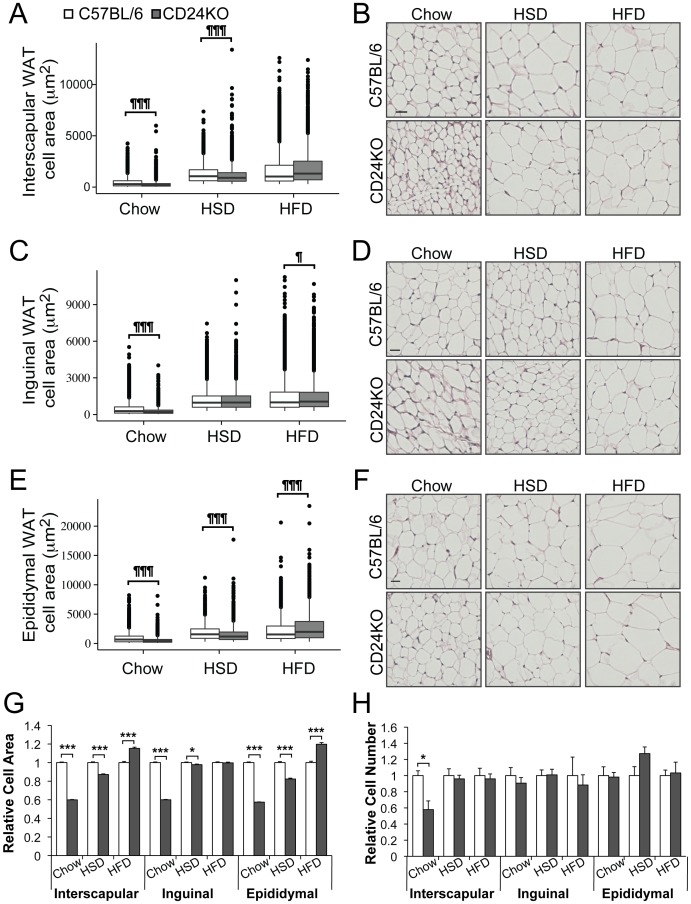
CD24 is a glycophosphatidylinositol (GPI)-linked cell surface receptor that is involved in regulating the survival or differentiation of several different cell types. CD24 has been used to identify pre-adipocytes that are able to reconstitute white adipose tissue (WAT) in vivo. Moreover, we recently found that the dynamic upregulation of CD24 in vitro during early phases of adipogenesis is necessary for mature adipocyte development. To determine the role of CD24 in adipocyte development in vivo, we evaluated the development of the inguinal and interscapular subcutaneous WAT and the epididymal visceral WAT in mice with a homozygous deletion of CD24 (CD24KO). We observed a significant decrease in WAT mass of 40% to 74% in WAT mass from both visceral and subcutaneous depots in male mice, with no significant effect in female mice, compared to wild-type (WT) sex- and age-matched controls. We also found that CD24KO mice had increased fasting glucose and free fatty acids, decreased fasting insulin, and plasma leptin. No major differences were observed in the sensitivity to insulin or glucose, or in circulating triglycerides, total cholesterol, HDL-cholesterol, or LDL-cholesterol levels between WT and CD24KO mice. Challenging the CD24KO mice with either high sucrose (35%) or high fat (45%) diets that promote increased adiposity, increased WAT mass and fasting insulin, adiponectin and leptin levels, as well as reduced the sensitivity to insulin and glucose, to the levels of WT mice on the same diets. The CD24-mediated reduction in fat pad size was due to a reduction in adipocyte cell size in all depots with no significant reduction pre-adipocyte or adipocyte cell number. Thus, we have clearly demonstrated that the global absence of CD24 affects adipocyte cell size in vivo in a sex- and diet-dependent manner, as well as causing metabolic disturbances in glucose homeostasis and free fatty acid levels.
Conflict of interest statement
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
