

Milkweed butterfly resistance to plant toxins is linked to sequestration, not coping with a toxic diet
- PMID: 26538594
- PMCID: PMC4650158
- DOI: 10.1098/rspb.2015.1865
Milkweed butterfly resistance to plant toxins is linked to sequestration, not coping with a toxic diet
Abstract
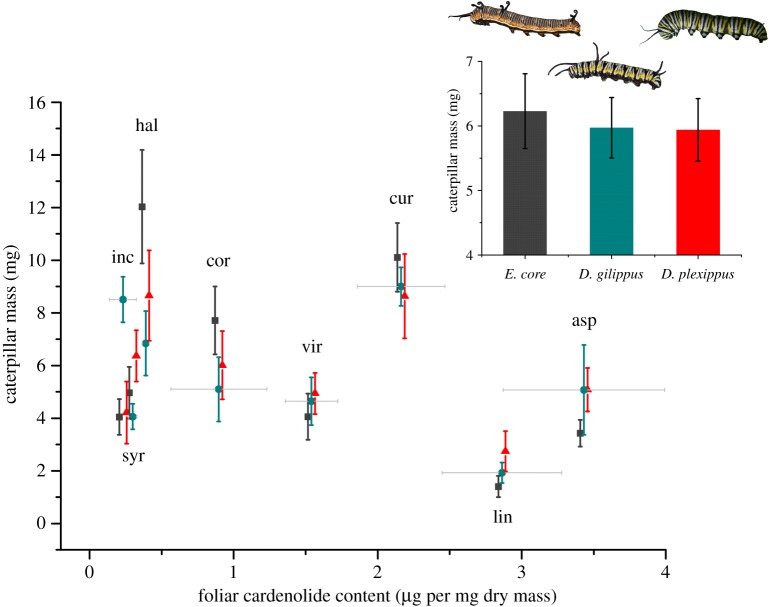
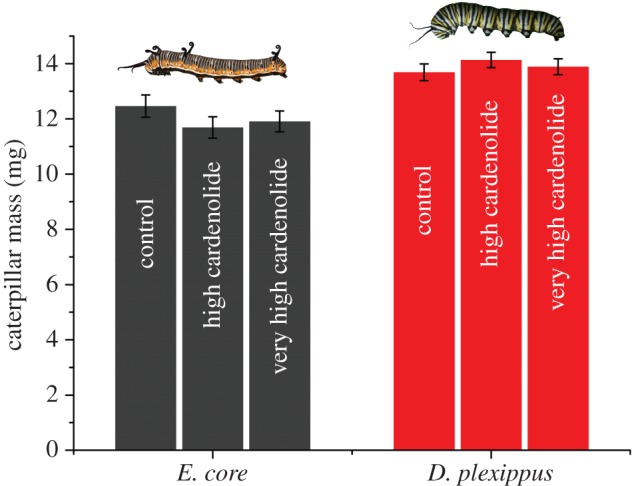
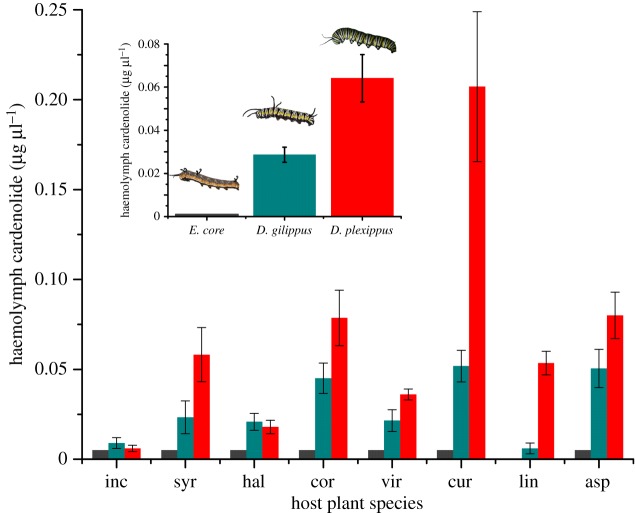
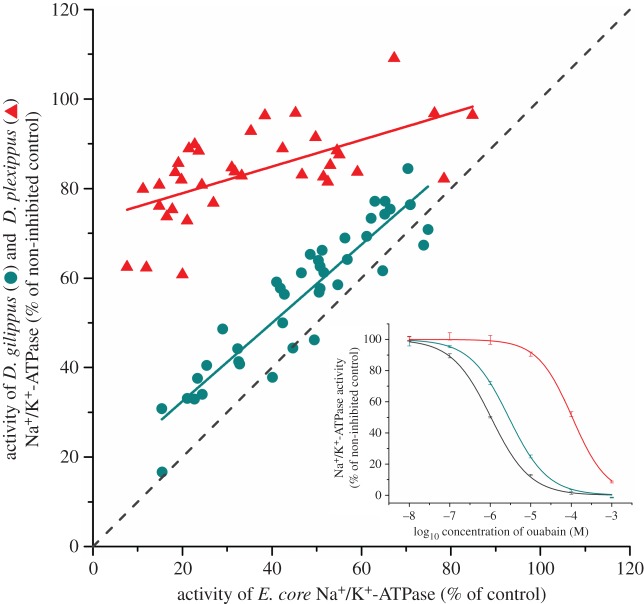
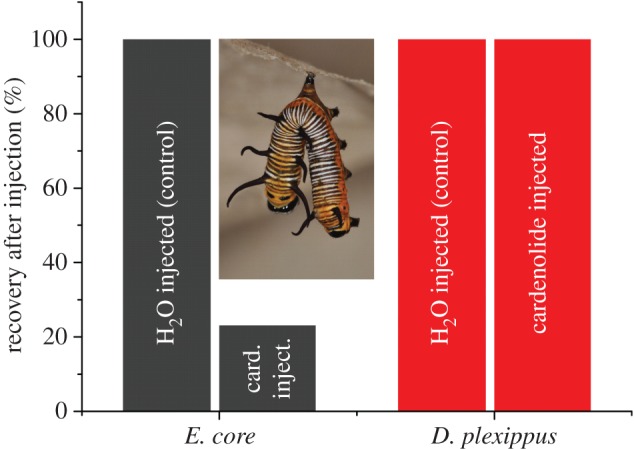
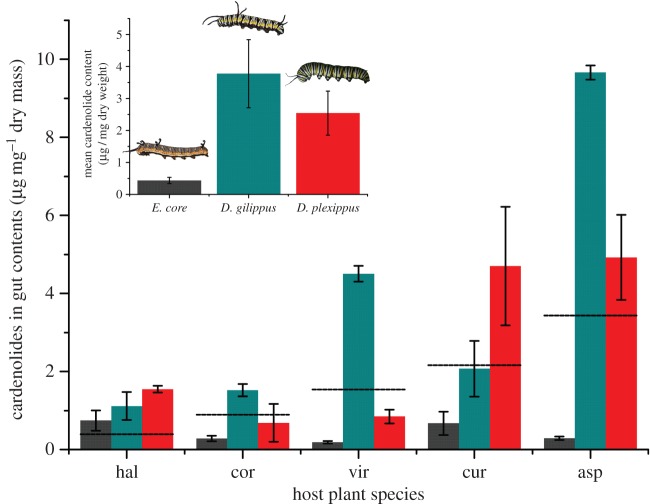
Insect resistance to plant toxins is widely assumed to have evolved in response to using defended plants as a dietary resource. We tested this hypothesis in the milkweed butterflies (Danaini) which have progressively evolved higher levels of resistance to cardenolide toxins based on amino acid substitutions of their cellular sodium-potassium pump (Na(+)/K(+)-ATPase). Using chemical, physiological and caterpillar growth assays on diverse milkweeds (Asclepias spp.) and isolated cardenolides, we show that resistant Na(+)/K(+)-ATPases are not necessary to cope with dietary cardenolides. By contrast, sequestration of cardenolides in the body (as a defence against predators) is associated with the three levels of Na(+)/K(+)-ATPase resistance. To estimate the potential physiological burden of cardenolide sequestration without Na(+)/K(+)-ATPase adaptations, we applied haemolymph of sequestering species on isolated Na(+)/K(+)-ATPase of sequestering and non-sequestering species. Haemolymph cardenolides dramatically impair non-adapted Na(+)/K(+)-ATPase, but had systematically reduced effects on Na(+)/K(+)-ATPase of sequestering species. Our data indicate that major adaptations to plant toxins may be evolutionarily linked to sequestration, and may not necessarily be a means to eat toxic plants. Na(+)/K(+)-ATPase adaptations thus were a potential mechanism through which predators spurred the coevolutionary arms race between plants and insects.
Keywords: adaptation; cardenolide; coevolution; milkweed butterflies; sequestration; tritrophic interaction.
© 2015 The Author(s).
Figures
References
-
- Ehrlich PR, Raven PH. 1964. Butterflies and plants: a study in coevolution. Evolution 18, 586–608. (10.2307/2406212) - DOI
-
- van Valen L. 1973. A new evolutionary law. Evol. Theory 1, 1–30.
-
- Singer MS, Stireman JO. 2005. The tri-trophic niche concept and adaptive radiation of phytophagous insects. Ecol. Lett. 8, 1247–1255. (10.1111/j.1461-0248.2005.00835.x) - DOI
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous