

Ion dynamics during seizures
- PMID: 26539081
- PMCID: PMC4612498
- DOI: 10.3389/fncel.2015.00419
Ion dynamics during seizures
Abstract
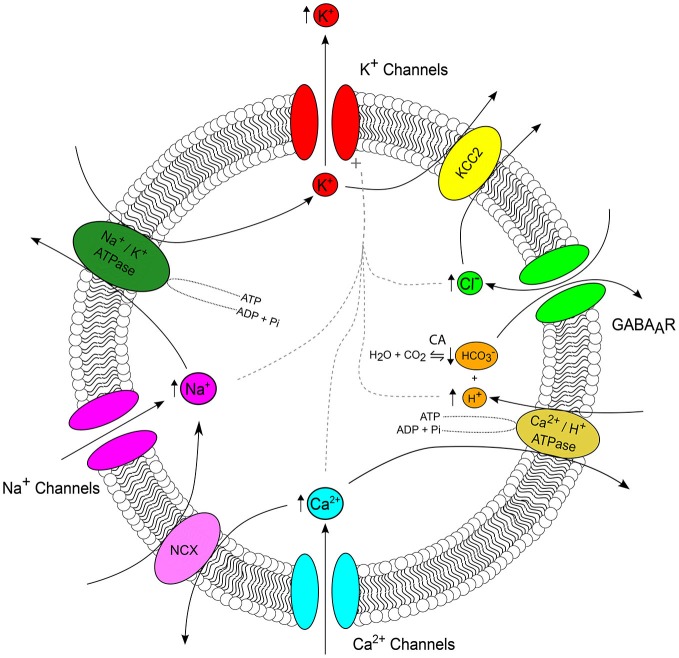
Changes in membrane voltage brought about by ion fluxes through voltage and transmitter-gated channels represent the basis of neural activity. As such, electrochemical gradients across the membrane determine the direction and driving force for the flow of ions and are therefore crucial in setting the properties of synaptic transmission and signal propagation. Ion concentration gradients are established by a variety of mechanisms, including specialized transporter proteins. However, transmembrane gradients can be affected by ionic fluxes through channels during periods of elevated neural activity, which in turn are predicted to influence the properties of on-going synaptic transmission. Such activity-induced changes to ion concentration gradients are a feature of both physiological and pathological neural processes. An epileptic seizure is an example of severely perturbed neural activity, which is accompanied by pronounced changes in intracellular and extracellular ion concentrations. Appreciating the factors that contribute to these ion dynamics is critical if we are to understand how a seizure event evolves and is sustained and terminated by neural tissue. Indeed, this issue is of significant clinical importance as status epilepticus-a type of seizure that does not stop of its own accord-is a life-threatening medical emergency. In this review we explore how the transmembrane concentration gradient of the six major ions (K(+), Na(+), Cl(-), Ca(2+), H(+)and [Formula: see text]) is altered during an epileptic seizure. We will first examine each ion individually, before describing how multiple interacting mechanisms between ions might contribute to concentration changes and whether these act to prolong or terminate epileptic activity. In doing so, we will consider how the availability of experimental techniques has both advanced and restricted our ability to study these phenomena.
Keywords: calcium; chloride; epilepsy; ion dynamics; pH; potassium; seizures; sodium.
Figures

References
-
- Avoli M., Louvel J., Pumain R., Olivier A. (1987). Seizure-like discharges induced by lowering [Mg2+]o in the human epileptogenic neocortex maintained in vitro. Brain Res. 417, 199–203. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous