

Aβ1-25-Derived Sphingolipid-Domain Tracer Peptide SBD Interacts with Membrane Ganglioside Clusters via a Coil-Helix-Coil Motif
- PMID: 26540054
- PMCID: PMC4661814
- DOI: 10.3390/ijms161125955
Aβ1-25-Derived Sphingolipid-Domain Tracer Peptide SBD Interacts with Membrane Ganglioside Clusters via a Coil-Helix-Coil Motif
Abstract
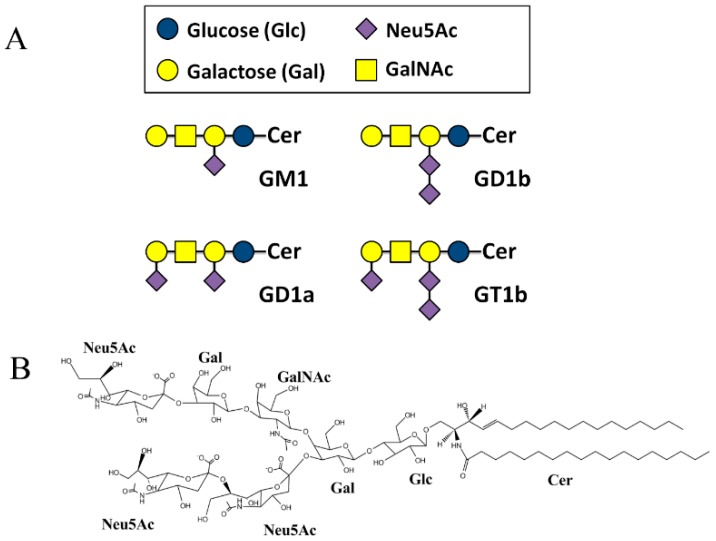
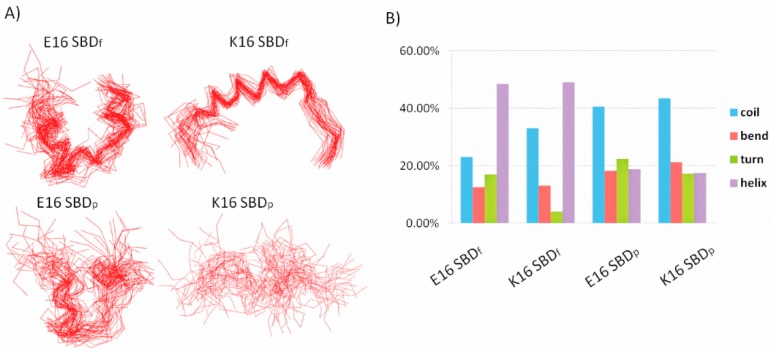
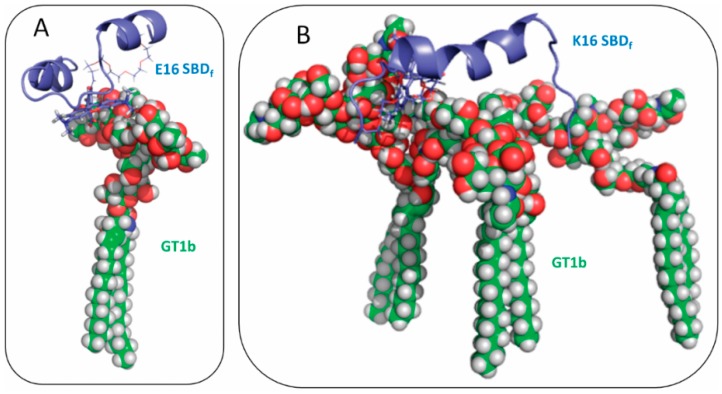
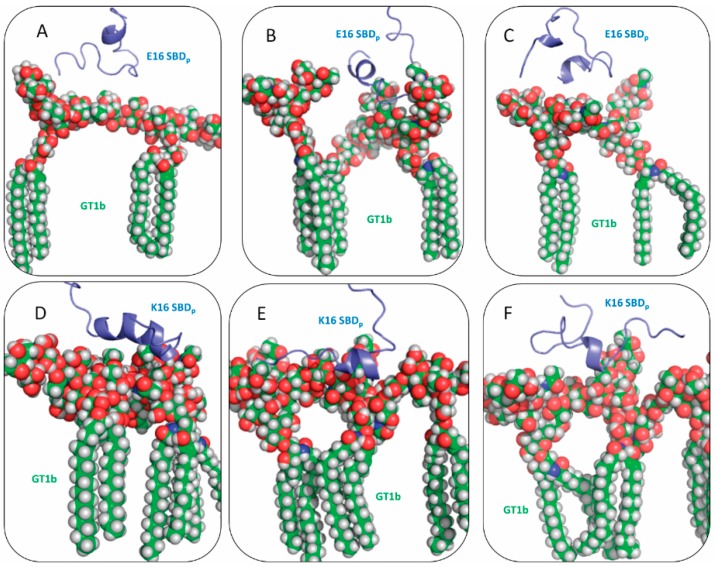
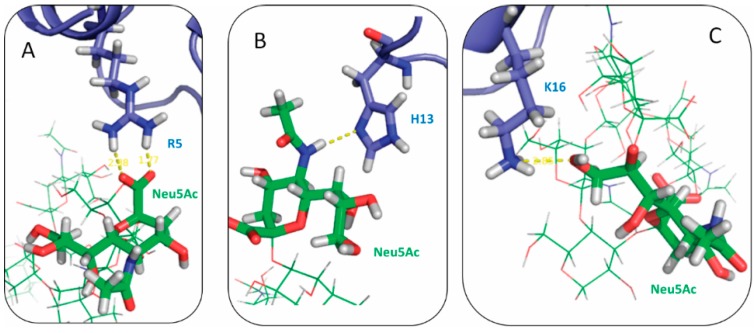
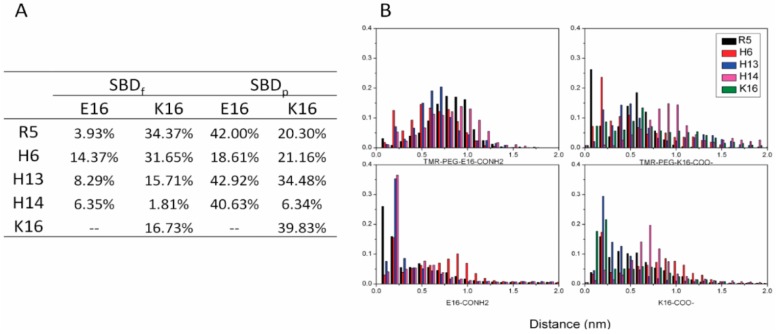
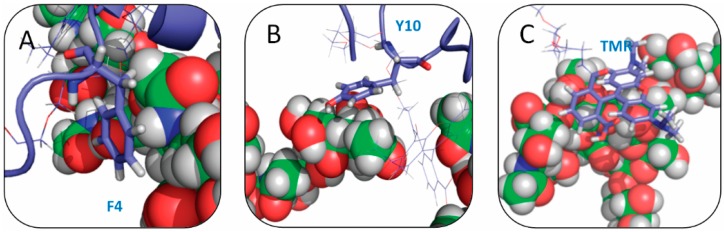
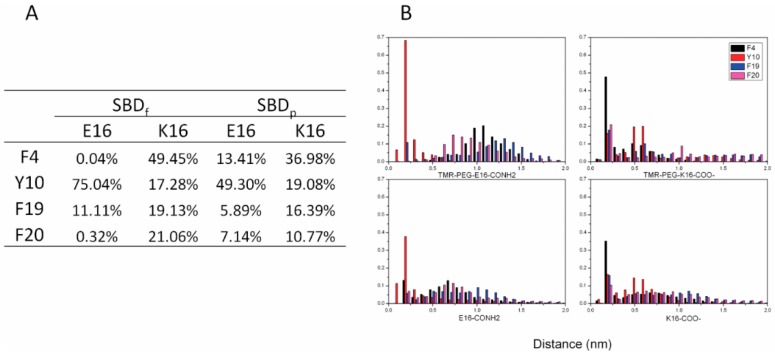
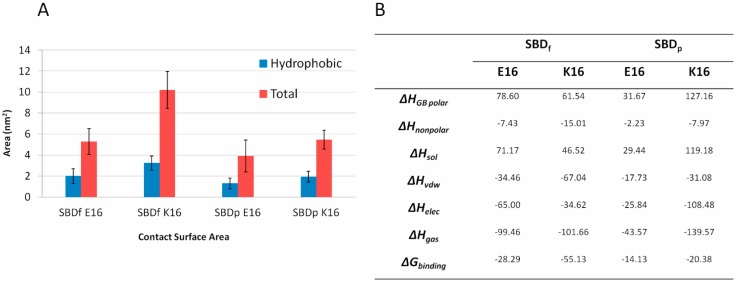
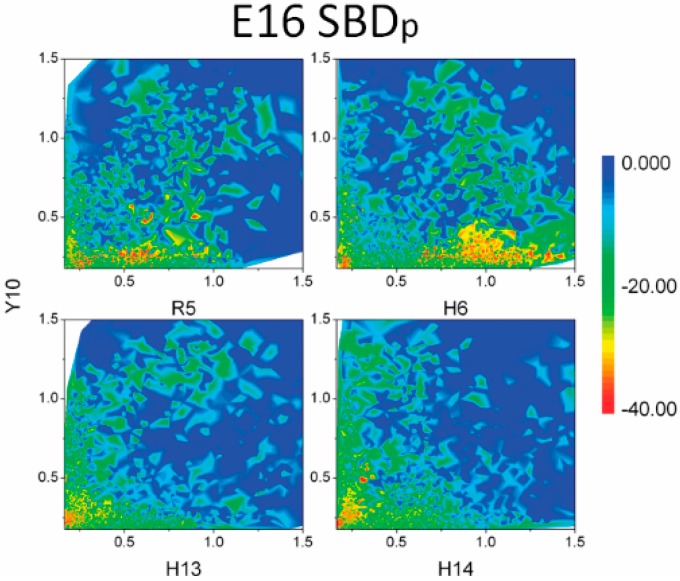
The Amyloid-β (Aβ)-derived, sphingolipid binding domain (SBD) peptide is a fluorescently tagged probe used to trace the diffusion behavior of sphingolipid-containing microdomains in cell membranes through binding to a constellation of glycosphingolipids, sphingomyelin, and cholesterol. However, the molecular details of the binding mechanism between SBD and plasma membrane domains remain unclear. Here, to investigate how the peptide recognizes the lipid surface at an atomically detailed level, SBD peptides in the environment of raft-like bilayers were examined in micro-seconds-long molecular dynamics simulations. We found that SBD adopted a coil-helix-coil structural motif, which binds to multiple GT1b gangliosides via salt bridges and CH-π interactions. Our simulation results demonstrate that the CH-π and electrostatic forces between SBD monomers and GT1b gangliosides clusters are the main driving forces in the binding process. The presence of the fluorescent dye and linker molecules do not change the binding mechanism of SBD probes with gangliosides, which involves the helix-turn-helix structural motif that was suggested to constitute a glycolipid binding domain common to some sphingolipid interacting proteins, including HIV gp120, prion, and Aβ.
Keywords: Sphingolipid binding domain; lipid rafts; molecular dynamics simulation.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
