

Cecropins from Plutella xylostella and Their Interaction with Metarhizium anisopliae
- PMID: 26544076
- PMCID: PMC4636316
- DOI: 10.1371/journal.pone.0142451
Cecropins from Plutella xylostella and Their Interaction with Metarhizium anisopliae
Abstract
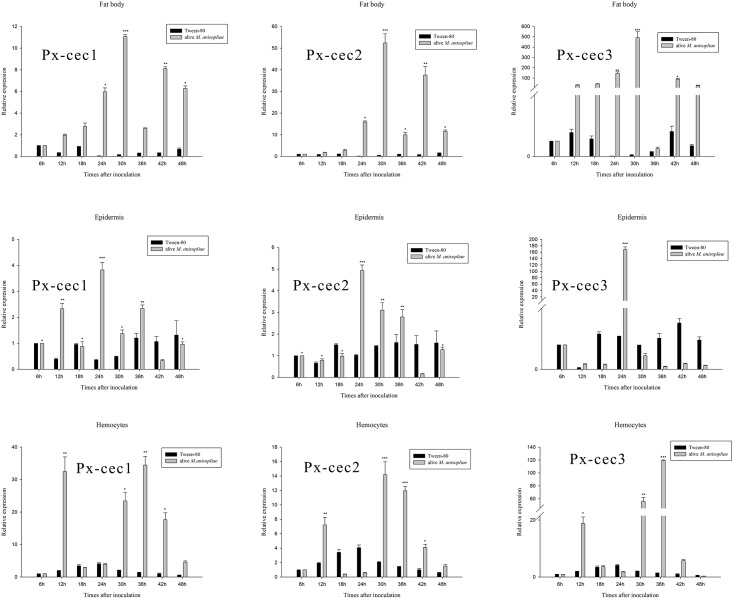
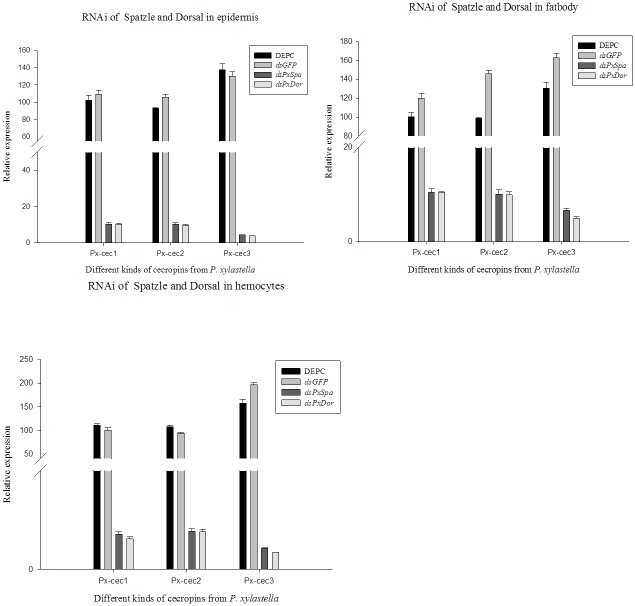
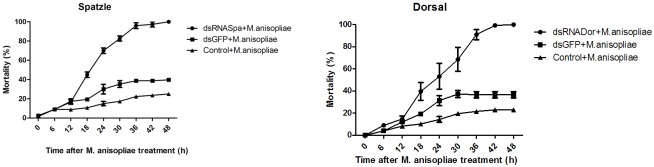
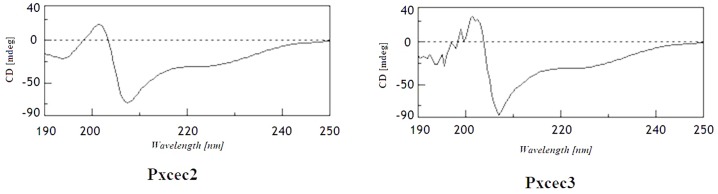
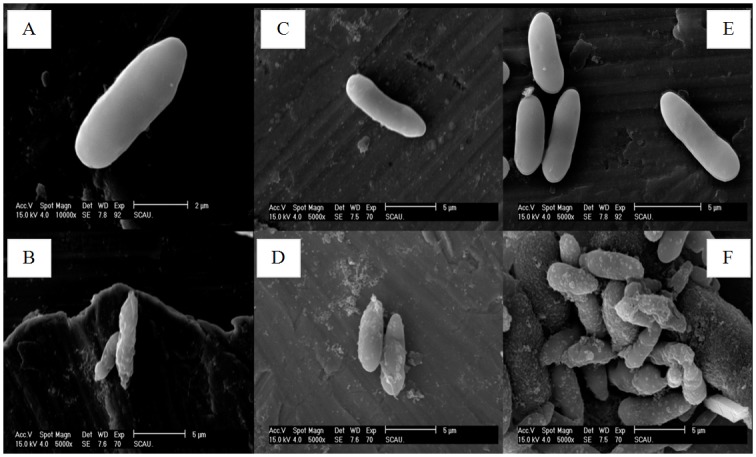
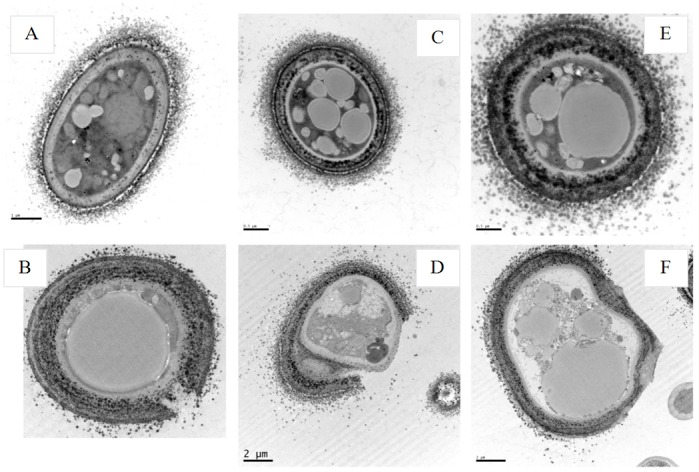
Cecropins are the most potent induced peptides to resist invading microorganisms. In the present study, two full length cDNA encoding cecropin2 (Px-cec2) and cecropin3 (Px-cec3) were obtained from P. xylostella by integrated analysis of genome and transcriptome data. qRT-PCR analysis revealed the high levels of transcripts of Px-cecs (Px-cec1, Px-cec2 and Px-cec3) in epidermis, fat body and hemocytes after 24, 30 and 36 h induction of Metarhizium anisopliae, respectively. Silencing of Spätzle and Dorsal separately caused the low expression of cecropins in the fat body, epidermis and hemocytes, and made the P.xylostella larvae more susceptible to M. anisopliae. Antimicrobial assays demonstrated that the purified recombinant cecropins, i.e., Px-cec1, Px-cec2 and Px-cec3, exerted a broad spectrum of antimicrobial activity against fungi, as well as Gram-positive and Gram-negative bacteria. Especially, Px-cecs showed higher activity against M. anisopliae than another selected fungi isolates. Scanning electron microscopy (SEM) and transmission electron microscopy (TEM) revealed that cecropins exerted the vital morphological alterations to the spores of M. anisopliae. Based on our results, cecropins played an imperative role in resisting infection of M. anisopliae, which will provide the foundation of biological control of insect pests by using cecorpins as a target in the future.
Conflict of interest statement
Figures

Similar articles
-
cDNA cloning and characterization of the antibacterial peptide cecropin 1 from the diamondback moth, Plutella xylostella L.Protein Expr Purif. 2012 Oct;85(2):230-8. doi: 10.1016/j.pep.2012.08.006. Epub 2012 Aug 17. Protein Expr Purif. 2012. PMID: 22921836
-
Studies on the interactions of neutral Galleria mellonella cecropin D with living bacterial cells.Amino Acids. 2019 Feb;51(2):175-191. doi: 10.1007/s00726-018-2641-4. Epub 2018 Aug 30. Amino Acids. 2019. PMID: 30167962
-
Novel antimicrobial anionic cecropins from the spruce budworm feature a poly-L-aspartic acid C-terminus.Proteins. 2021 Sep;89(9):1205-1215. doi: 10.1002/prot.26142. Epub 2021 May 24. Proteins. 2021. PMID: 33973678
-
Molecular Identification of a Moricin Family Antimicrobial Peptide (Px-Mor) From Plutella xylostella With Activities Against the Opportunistic Human Pathogen Aureobasidium pullulans.Front Microbiol. 2019 Oct 11;10:2211. doi: 10.3389/fmicb.2019.02211. eCollection 2019. Front Microbiol. 2019. PMID: 31681182 Free PMC article.
-
Severe fungal sclerokeratitis caused by Metarhizium anisopliae: a case report and literature review.Mycoses. 2015 Feb;58(2):88-92. doi: 10.1111/myc.12279. Epub 2015 Jan 15. Mycoses. 2015. PMID: 25590990 Review.
Cited by
-
Characterization and Antimicrobial Activity of Amphiphilic Peptide AP3 and Derivative Sequences.Antibiotics (Basel). 2019 Mar 6;8(1):20. doi: 10.3390/antibiotics8010020. Antibiotics (Basel). 2019. PMID: 30845708 Free PMC article.
-
Insect Meals and Insect Antimicrobial Peptides as an Alternative for Antibiotics and Growth Promoters in Livestock Production.Pathogens. 2023 Jun 20;12(6):854. doi: 10.3390/pathogens12060854. Pathogens. 2023. PMID: 37375544 Free PMC article. Review.
-
Antimicrobial Peptides and Proteins: From Nature's Reservoir to the Laboratory and Beyond.Front Chem. 2021 Jun 18;9:691532. doi: 10.3389/fchem.2021.691532. eCollection 2021. Front Chem. 2021. PMID: 34222199 Free PMC article. Review.
-
Chemical Compounds and Bioactivity of Aqueous Extracts of Alibertia spp. in the Control of Plutella xylostella L. (Lepidoptera: Plutellidae).Insects. 2017 Nov 22;8(4):125. doi: 10.3390/insects8040125. Insects. 2017. PMID: 29165339 Free PMC article.
-
Insect Antimicrobial Peptides, a Mini Review.Toxins (Basel). 2018 Nov 8;10(11):461. doi: 10.3390/toxins10110461. Toxins (Basel). 2018. PMID: 30413046 Free PMC article. Review.
References
-
- Hultmark D.Drosophila immunity: paths and patterns. Current Opinion in Immunology. 2003; 15(1): 12–19. - PubMed
-
- Engel E, Viargues P, Mortier M, Taillebourg E, Couté Y. Identifying USPs regulating immune signals in Drosophila: USP2 deubiquitinates Imd and promotes its degradation by interacting with the proteasome. Cell Communication and Signaling.2014; 12: 41–43. 10.1186/s12964-014-0041-2 - DOI - PMC - PubMed
-
- Charroux B, Royet J. Drosophila immune response: From systemic antimicrobial peptide production in fat body cells to local defense in the intestinal tract. Fly.2014; 4(1):40–47. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
