

Single-Stranded DNA Cleavage by Divergent CRISPR-Cas9 Enzymes
- PMID: 26545076
- PMCID: PMC4636735
- DOI: 10.1016/j.molcel.2015.10.030
Single-Stranded DNA Cleavage by Divergent CRISPR-Cas9 Enzymes
Abstract
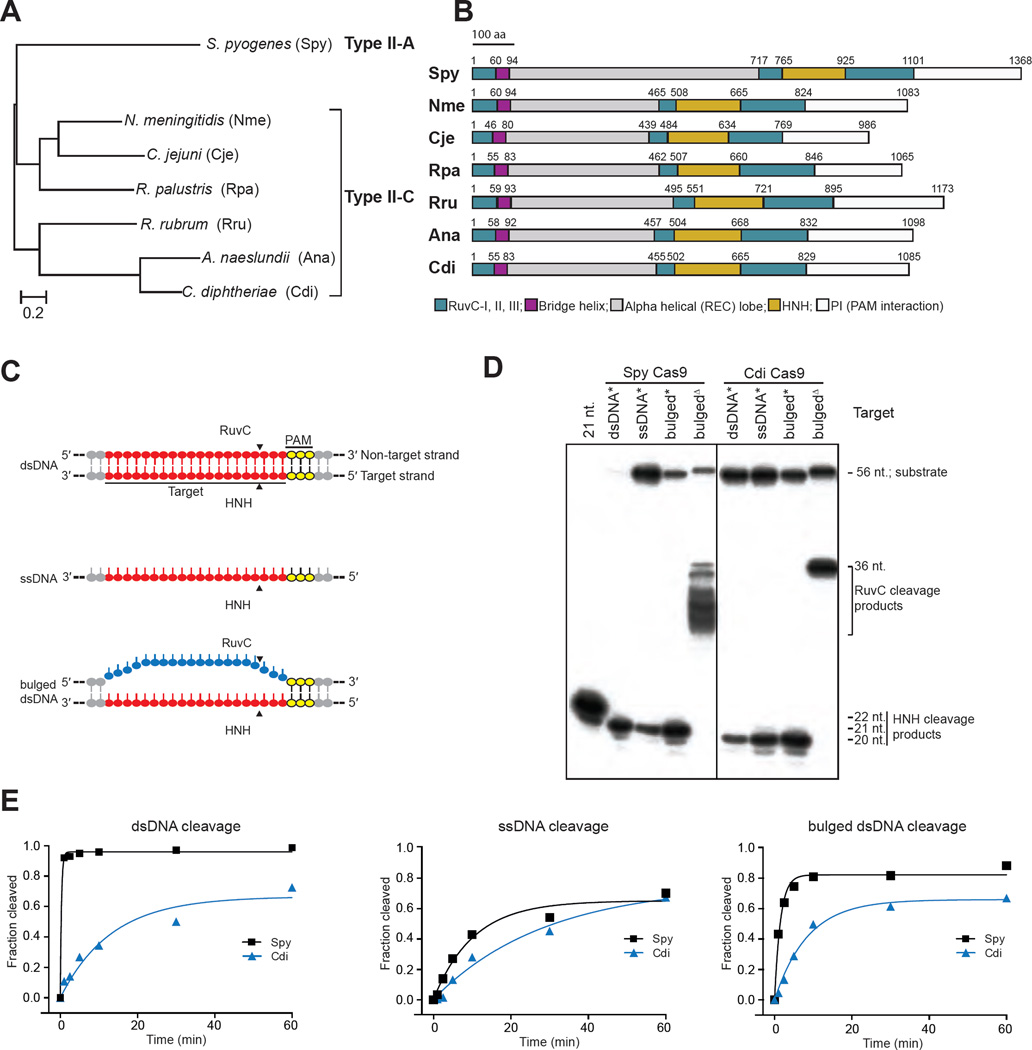
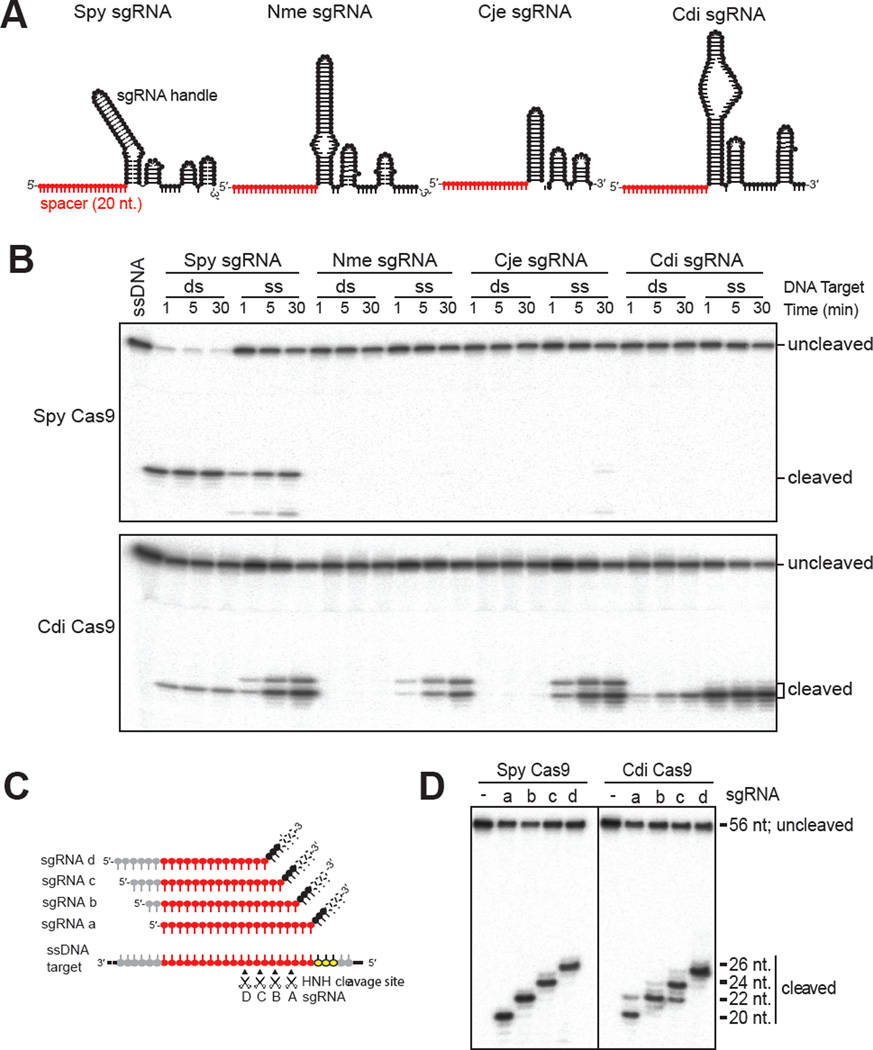
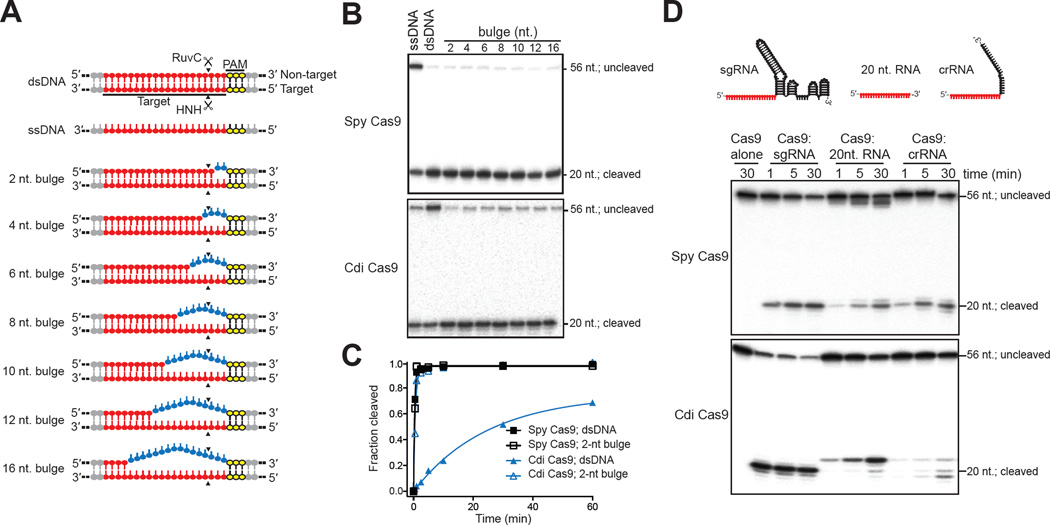
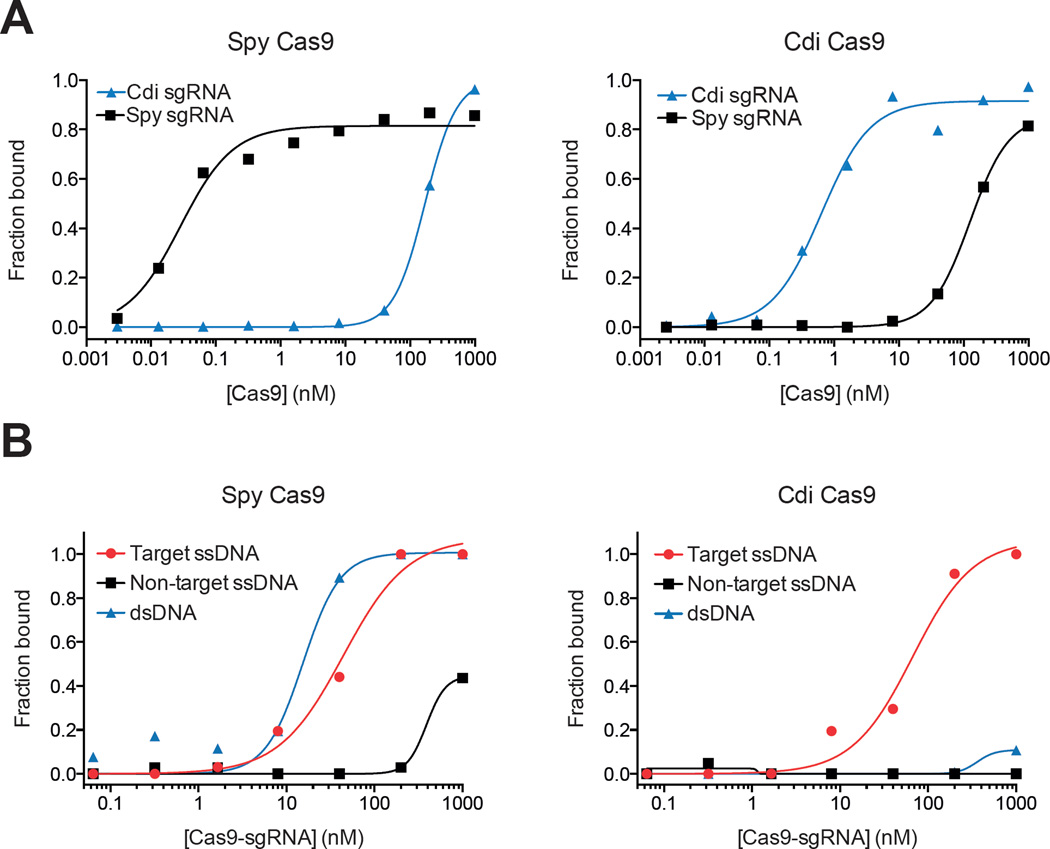
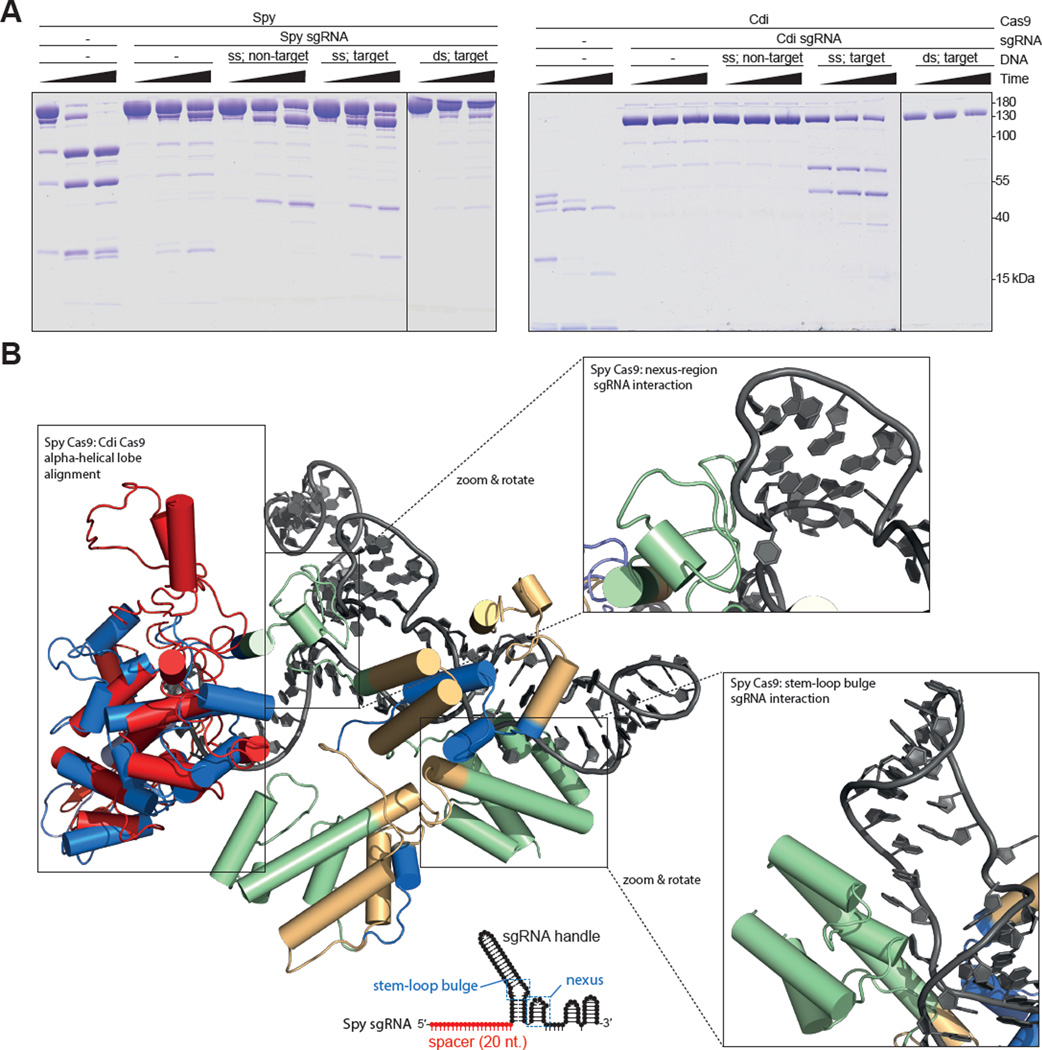
Double-stranded DNA (dsDNA) cleavage by Cas9 is a hallmark of type II CRISPR-Cas immune systems. Cas9-guide RNA complexes recognize 20-base-pair sequences in DNA and generate a site-specific double-strand break, a robust activity harnessed for genome editing. DNA recognition by all studied Cas9 enzymes requires a protospacer adjacent motif (PAM) next to the target site. We show that Cas9 enzymes from evolutionarily divergent bacteria can recognize and cleave single-stranded DNA (ssDNA) by an RNA-guided, PAM-independent recognition mechanism. Comparative analysis shows that in contrast to the type II-A S. pyogenes Cas9 that is widely used for genome engineering, the smaller type II-C Cas9 proteins have limited dsDNA binding and unwinding activity and promiscuous guide RNA specificity. These results indicate that inefficiency of type II-C Cas9 enzymes for genome editing results from a limited ability to cleave dsDNA and suggest that ssDNA cleavage was an ancestral function of the Cas9 enzyme family.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Briner AE, Donohoue PD, Gomaa AA, Selle K, Slorach EM, Nye CH, Haurwitz RE, Beisel CL, May AP, Barrangou R. Guide RNA Functional Modules Direct Cas9 Activity and Orthogonality. Mol. Cell. 2014;56:333–339. - PubMed
-
- Chase JW, Williams KR. Single-stranded DNA binding proteins required for DNA replication. Annu. Rev. Biochem. 1986;55:103–136. - PubMed
-
- Deng L, Garrett RA, Shah SA, Peng X, She Q. A novel interference mechanism by a type IIIB CRISPR-Cmr module in Sulfolobus. Mol. Microbiol. 2013;87:1088–1099. - PubMed
-
- Doudna JA, Charpentier E. The new frontier of genome engineering with CRISPR-Cas9. Sci. 2014;346 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
