

Novel N-terminal and Lysine Methyltransferases That Target Translation Elongation Factor 1A in Yeast and Human
- PMID: 26545399
- PMCID: PMC4762513
- DOI: 10.1074/mcp.M115.052449
Novel N-terminal and Lysine Methyltransferases That Target Translation Elongation Factor 1A in Yeast and Human
Abstract
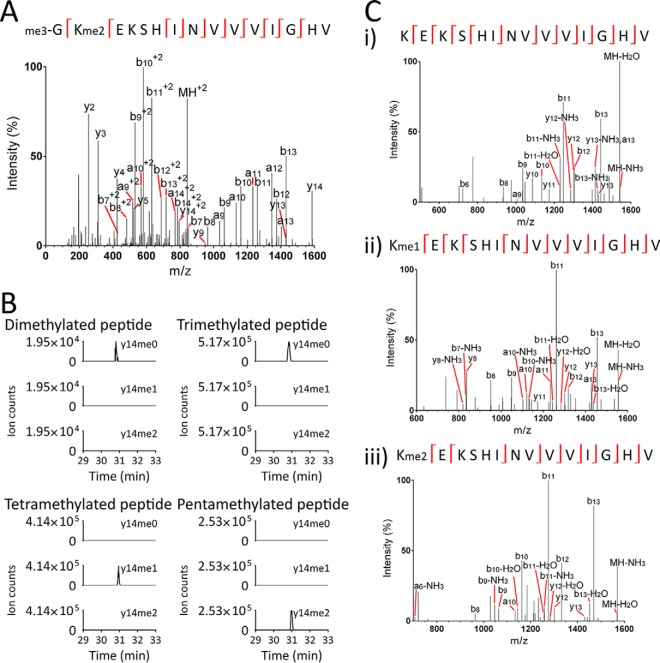
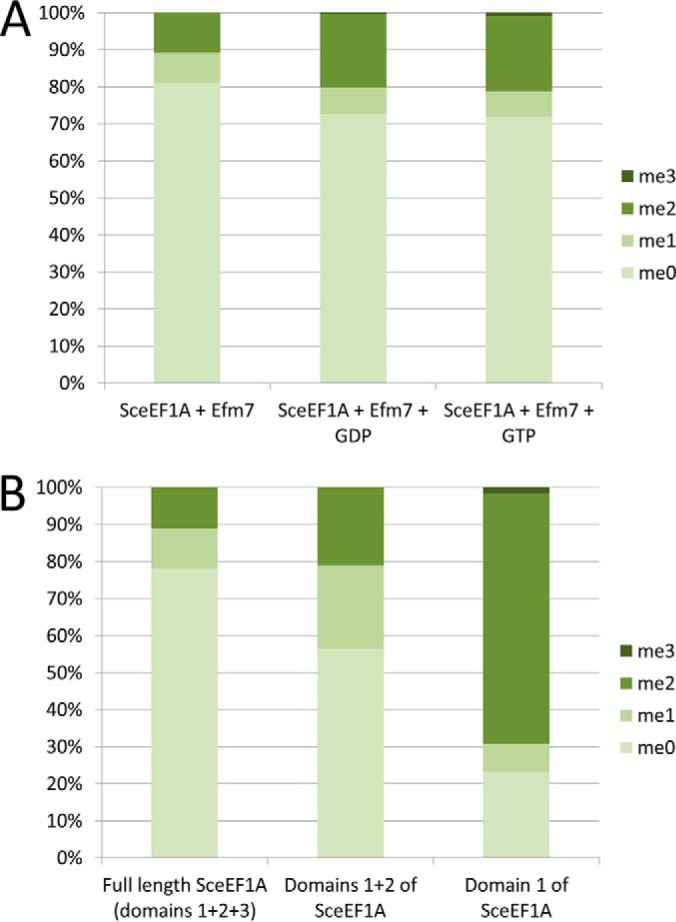
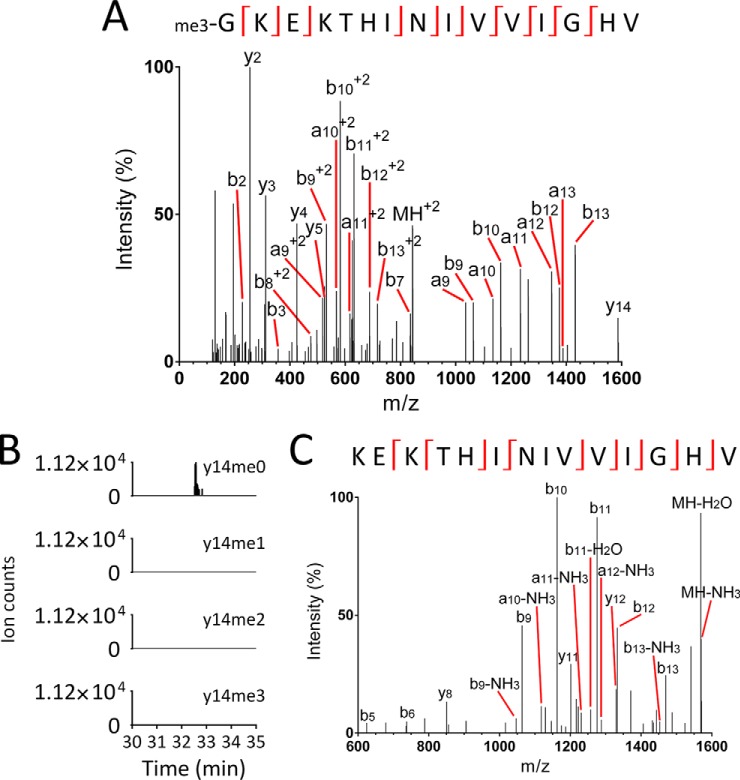
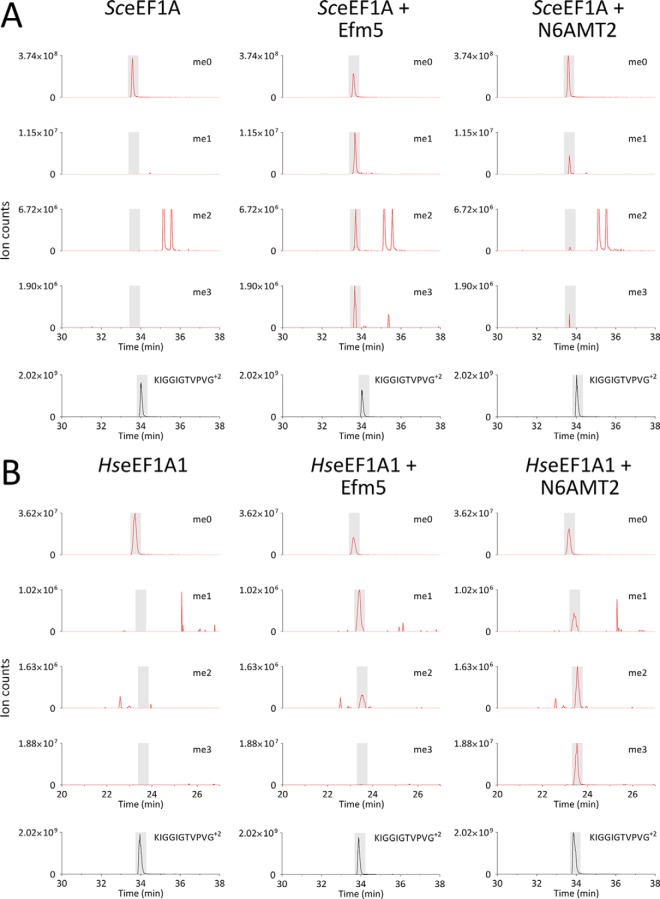
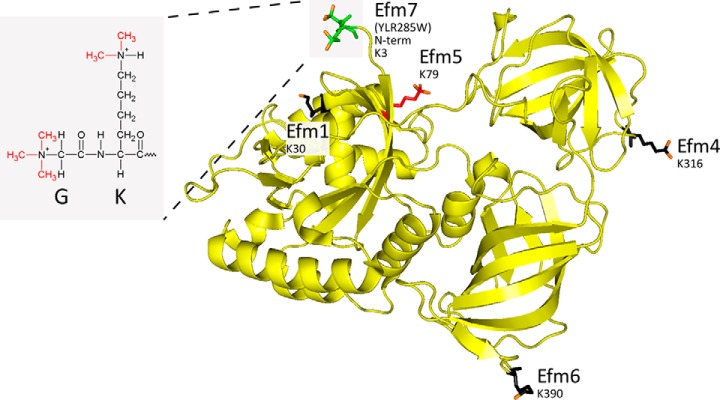
Eukaryotic elongation factor 1A (eEF1A) is an essential, highly methylated protein that facilitates translational elongation by delivering aminoacyl-tRNAs to ribosomes. Here, we report a new eukaryotic protein N-terminal methyltransferase, Saccharomyces cerevisiae YLR285W, which methylates eEF1A at a previously undescribed high-stoichiometry N-terminal site and the adjacent lysine. Deletion of YLR285W resulted in the loss of N-terminal and lysine methylation in vivo, whereas overexpression of YLR285W resulted in an increase of methylation at these sites. This was confirmed by in vitro methylation of eEF1A by recombinant YLR285W. Accordingly, we name YLR285W as elongation factor methyltransferase 7 (Efm7). This enzyme is a new type of eukaryotic N-terminal methyltransferase as, unlike the three other known eukaryotic N-terminal methyltransferases, its substrate does not have an N-terminal [A/P/S]-P-K motif. We show that the N-terminal methylation of eEF1A is also present in human; this conservation over a large evolutionary distance suggests it to be of functional importance. This study also reports that the trimethylation of Lys(79) in eEF1A is conserved from yeast to human. The methyltransferase responsible for Lys(79) methylation of human eEF1A is shown to be N6AMT2, previously documented as a putative N(6)-adenine-specific DNA methyltransferase. It is the direct ortholog of the recently described yeast Efm5, and we show that Efm5 and N6AMT2 can methylate eEF1A from either species in vitro. We therefore rename N6AMT2 as eEF1A-KMT1. Including the present work, yeast eEF1A is now documented to be methylated by five different methyltransferases, making it one of the few eukaryotic proteins to be extensively methylated by independent enzymes. This implies more extensive regulation of eEF1A by this posttranslational modification than previously appreciated.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures


References
-
- Erce M. A., Pang C. N., Hart-Smith G., and Wilkins M. R. (2012) The methylproteome and the intracellular methylation network. Proteomics 12, 564–586 - PubMed
-
- Winter D. L., Erce M. A., and Wilkins M. R. (2014) A web of possibilities: Network-based discovery of protein interaction codes. J. Proteome Res. 13, 5333–5338 - PubMed
-
- Jenuwein T., and Allis C. D. (2001) Translating the histone code. Science 293, 1074–1080 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
