

Up-regulation of abscisic acid signaling pathway facilitates aphid xylem absorption and osmoregulation under drought stress
- PMID: 26546578
- PMCID: PMC4737068
- DOI: 10.1093/jxb/erv481
Up-regulation of abscisic acid signaling pathway facilitates aphid xylem absorption and osmoregulation under drought stress
Abstract
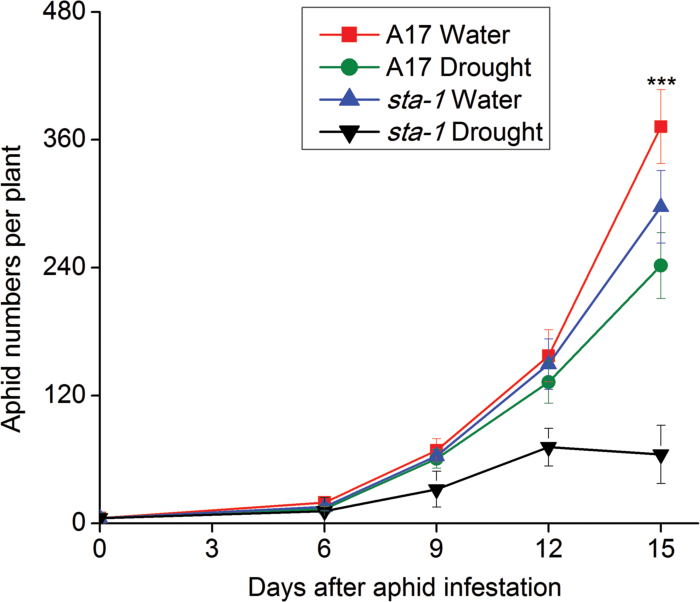
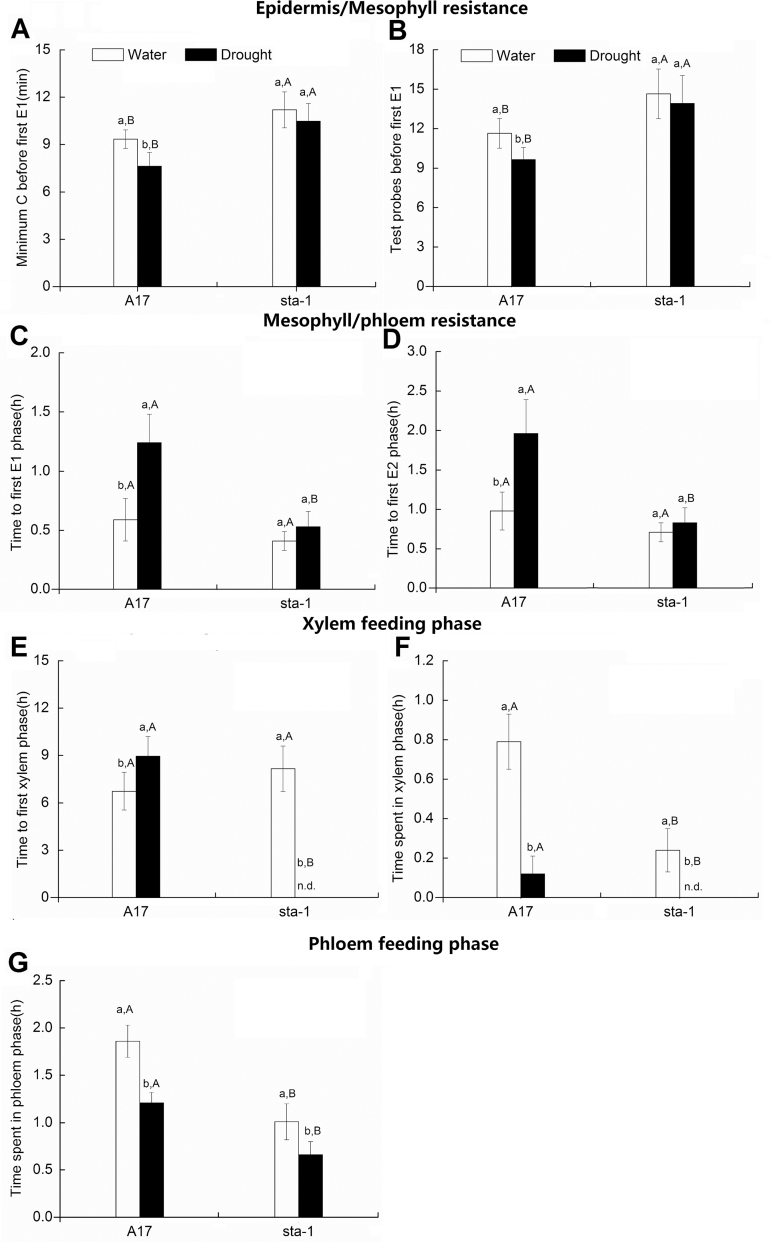
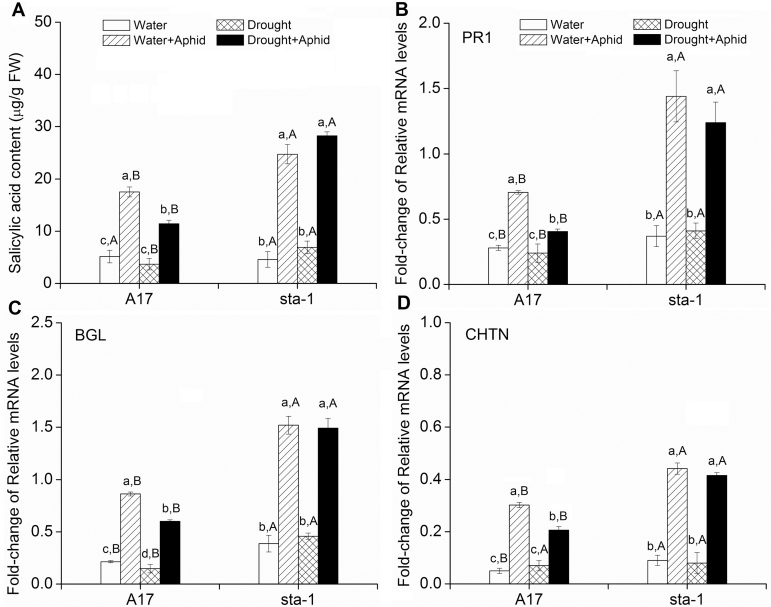
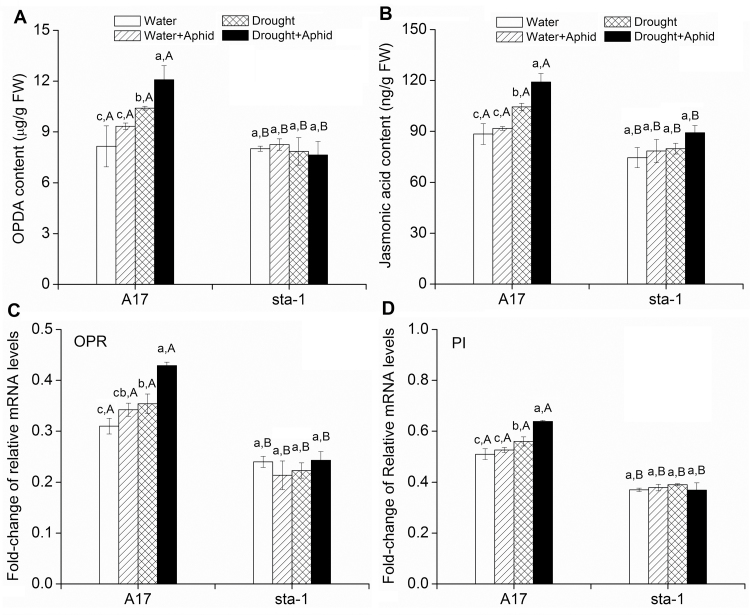
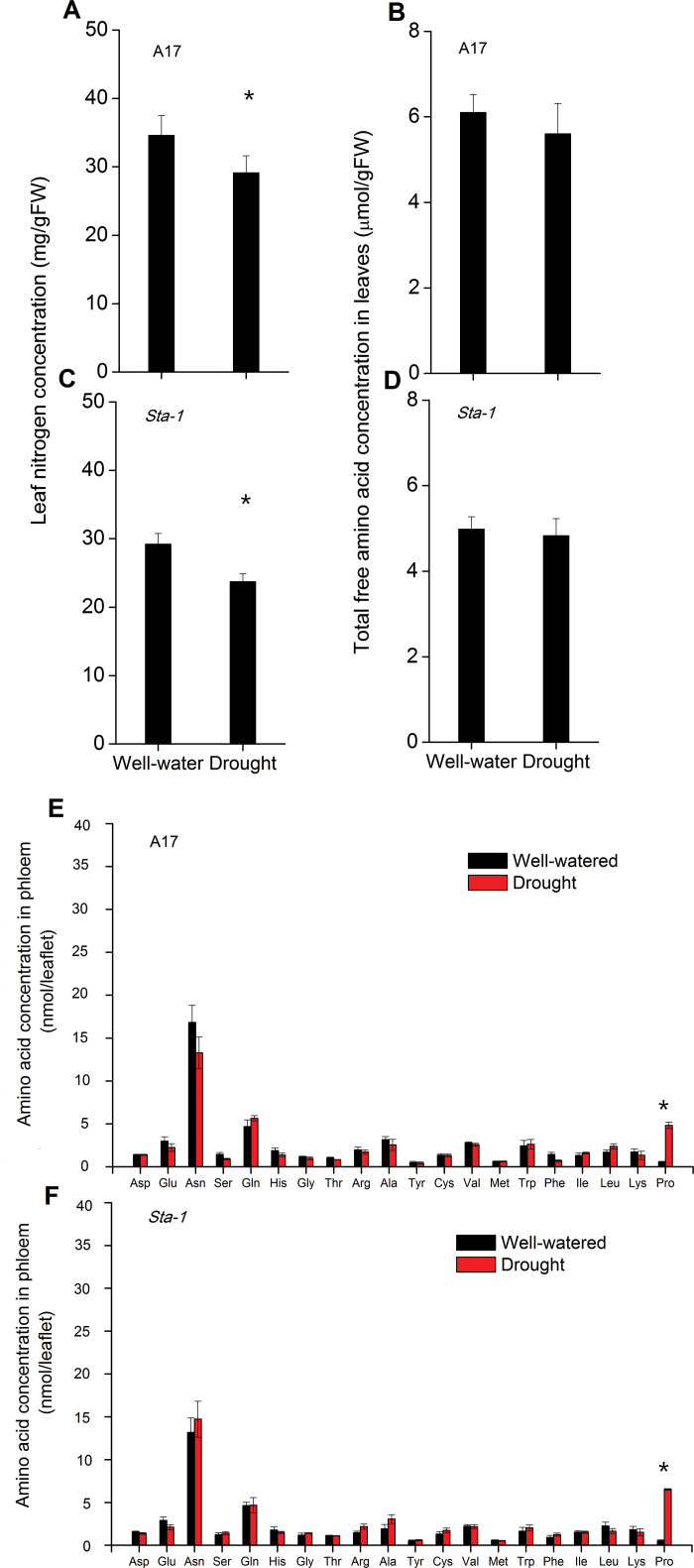
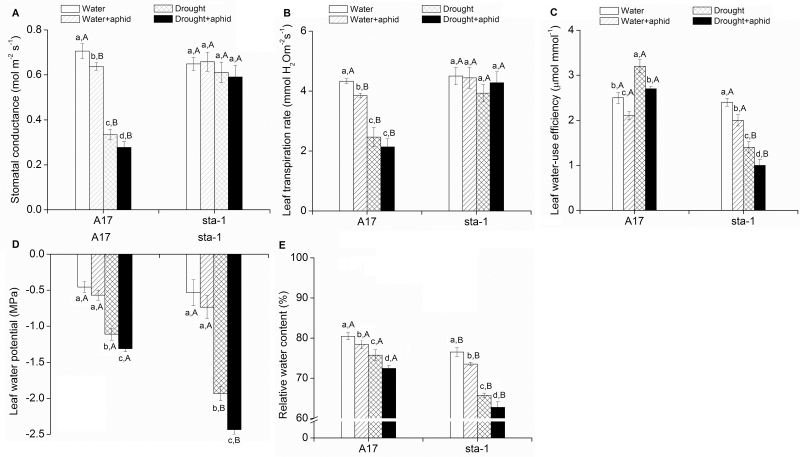
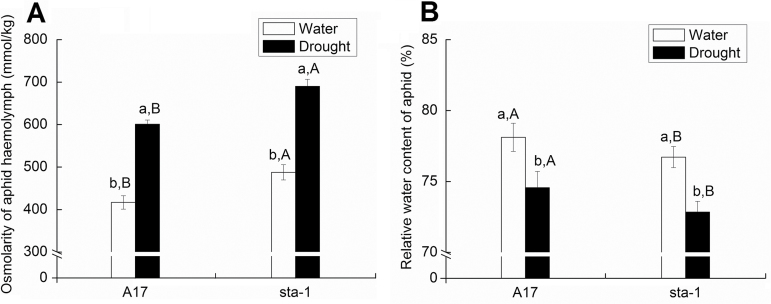
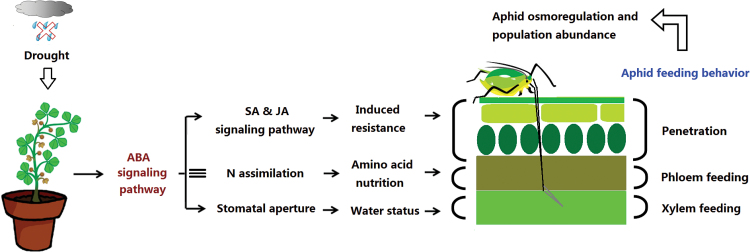
The activation of the abscisic acid (ABA) signaling pathway reduces water loss from plants challenged by drought stress. The effect of drought-induced ABA signaling on the defense and nutrition allocation of plants is largely unknown. We postulated that these changes can affect herbivorous insects. We studied the effects of drought on different feeding stages of pea aphids in the wild-type A17 of Medicago truncatula and ABA signaling pathway mutant sta-1. We examined the impact of drought on plant water status, induced plant defense signaling via the abscisic acid (ABA), jasmonic acid (JA), and salicylic acid (SA) pathways, and on the host nutritional quality in terms of leaf free amino acid content. During the penetration phase of aphid feeding, drought decreased epidermis/mesophyll resistance but increased mesophyll/phloem resistance of A17 but not sta-1 plants. Quantification of transcripts associated with ABA, JA and SA signaling indicated that the drought-induced up-regulation of ABA signaling decreased the SA-dependent defense but increased the JA-dependent defense in A17 plants. During the phloem-feeding phase, drought had little effect on the amino acid concentrations and the associated aphid phloem-feeding parameters in both plant genotypes. In the xylem absorption stage, drought decreased xylem absorption time of aphids in both genotypes because of decreased water potential. Nevertheless, the activation of the ABA signaling pathway increased water-use efficiency of A17 plants by decreasing the stomatal aperture and transpiration rate. In contrast, the water potential of sta-1 plants (unable to close stomata) was too low to support xylem absorption activity of aphids; the aphids on sta-1 plants had the highest hemolymph osmolarity and lowest abundance under drought conditions. Taken together this study illustrates the significance of cross-talk between biotic-abiotic signaling pathways in plant-aphid interaction, and reveals the mechanisms leading to alter aphid fecundity in water stresses plants.
Keywords: Abscisic acid; Acyrthosiphon pisum; Medicago truncatula; drought; feeding behavior; jasmonic acid; osmoregulation; resistance; salicylic acid; xylem absorption..
© The Author 2015. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures

References
-
- Alvarez AE, Tjallingii WF, Garzo E, Vleeshouwers V, Dicke M, Vosman B. 2006. Location of resistance factors in the leaves of potato and wild tuber-bearing Solanum species to the aphid Myzus persicae . Entomologia Experimentalis et Applicata 121, 145–157.
-
- Asselbergh B, Curvers K, França SC, Audenaert K, Vuylsteke M, Van Breusegem F, Höfte M. 2007. Resistance to Botrytis cinerea in sitiens, an abscisic acid-deficient tomato mutant, involves timely production of hydrogen peroxide and cell wall modifications in the epidermis. Plant Physiology 144, 1863–1877. - PMC - PubMed
-
- Barrs HD, Weatherley PE. 1962. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Australian Journal of Biological Sciences 15, 413–428.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
