

Acute resistance exercise activates rapamycin-sensitive and -insensitive mechanisms that control translational activity and capacity in skeletal muscle
- PMID: 26548696
- PMCID: PMC4713726
- DOI: 10.1113/JP271365
Acute resistance exercise activates rapamycin-sensitive and -insensitive mechanisms that control translational activity and capacity in skeletal muscle
Abstract
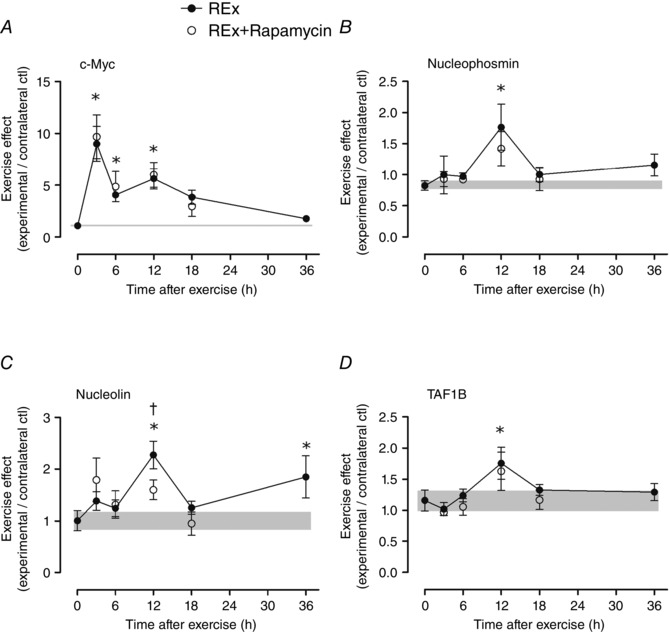
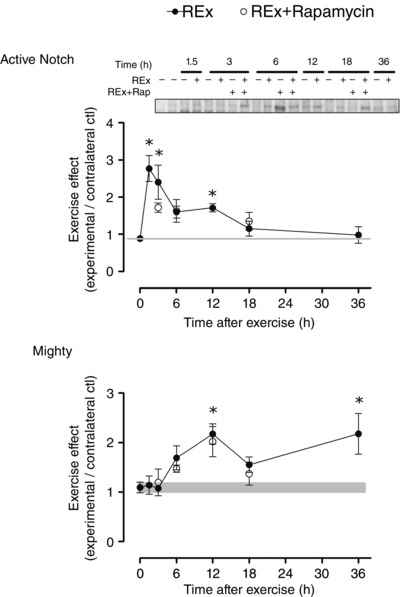
Key points: Ribosome biogenesis is the primary determinant of translational capacity, but its regulation in skeletal muscle following acute resistance exercise is poorly understood. Resistance exercise increases muscle protein synthesis acutely, and muscle mass with training, but the role of translational capacity in these processes is unclear. Here, we show that acute resistance exercise activated pathways controlling translational activity and capacity through both rapamycin-sensitive and -insensitive mechanisms. Transcription factor c-Myc and its downstream targets, which are known to regulate ribosome biogenesis in other cell types, were upregulated after resistance exercise in a rapamycin-independent manner and may play a role in determining translational capacity in skeletal muscle. Local inhibition of myostatin was also not affected by rapamycin and may contribute to the rapamycin-independent effects of resistance exercise.
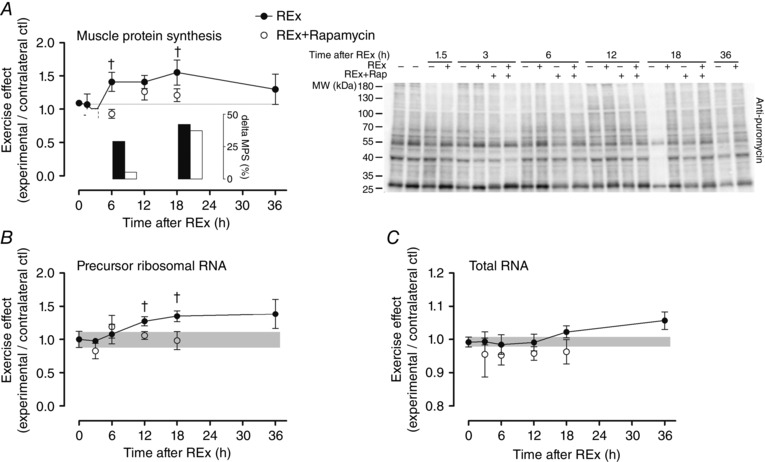
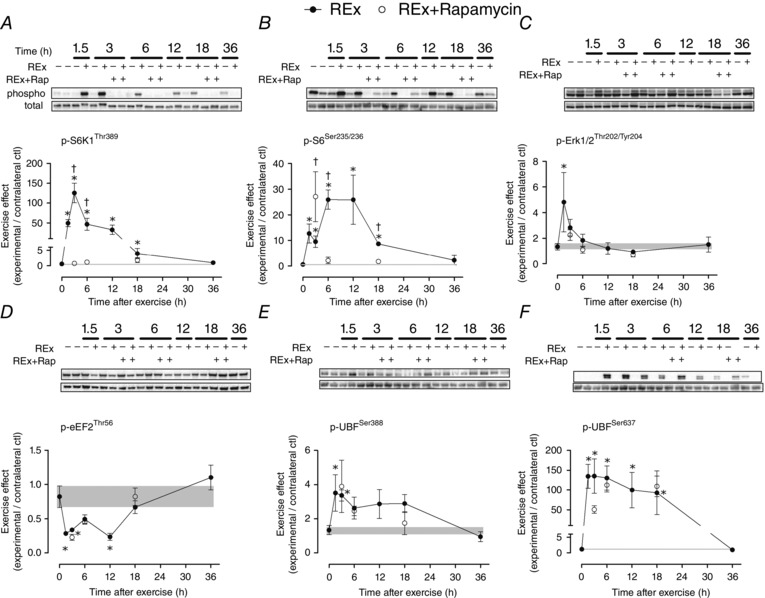
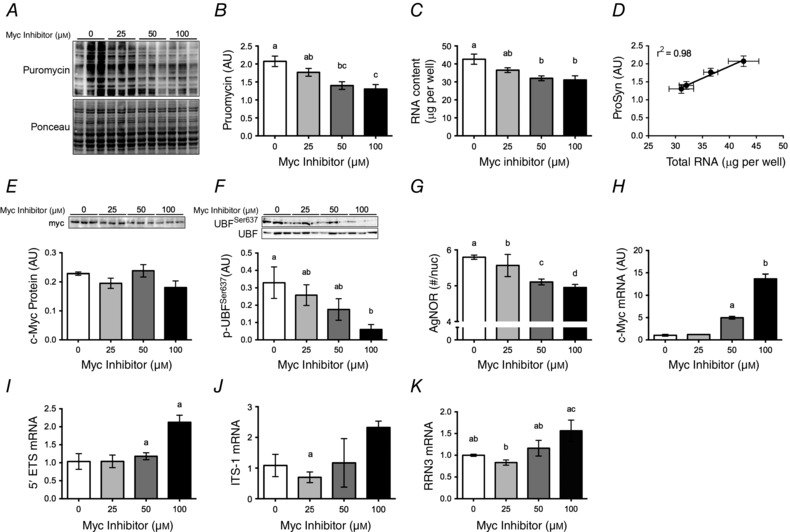
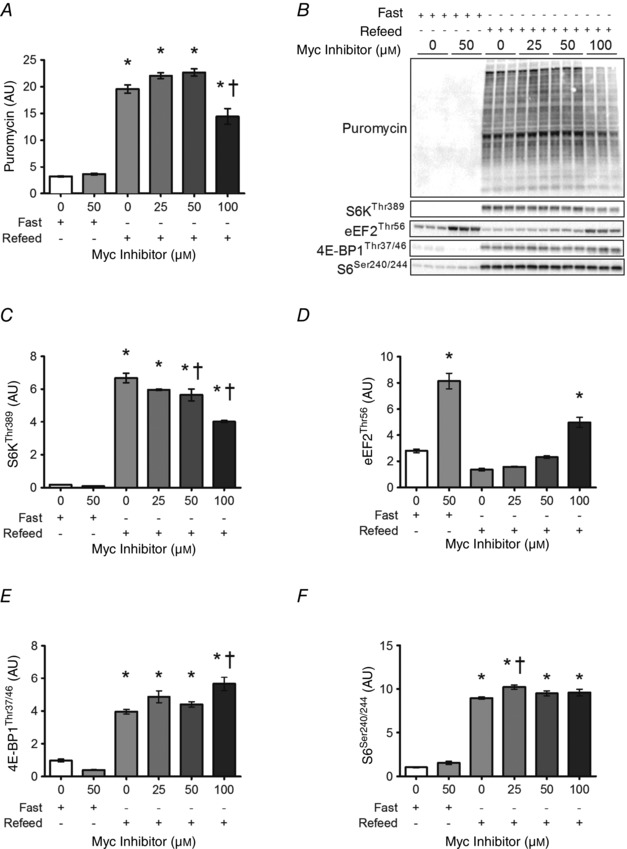
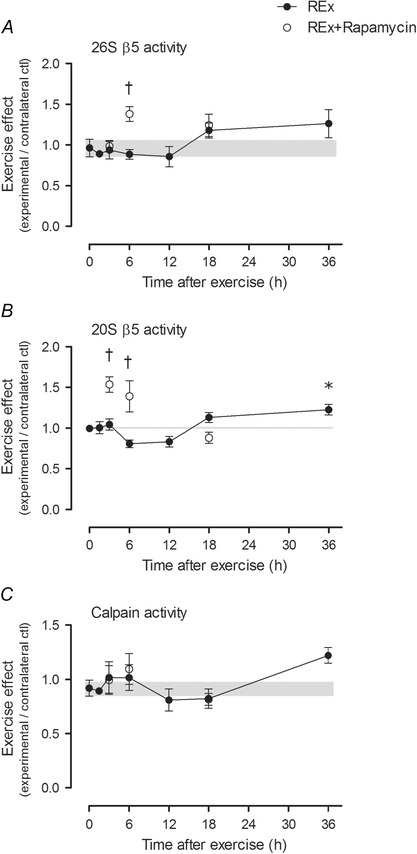
Abstract: This study aimed to determine (1) the effect of acute resistance exercise on mechanisms of ribosome biogenesis, and (2) the impact of mammalian target of rapamycin on ribosome biogenesis, and muscle protein synthesis (MPS) and degradation. Female F344BN rats underwent unilateral electrical stimulation of the sciatic nerve to mimic resistance exercise in the tibialis anterior (TA) muscle. TA muscles were collected at intervals over the 36 h of exercise recovery (REx); separate groups of animals were administered rapamycin pre-exercise (REx+Rapamycin). Resistance exercise led to a prolonged (6-36 h) elevation (30-50%) of MPS that was fully blocked by rapamycin at 6 h but only partially at 18 h. REx also altered pathways that regulate protein homeostasis and mRNA translation in a manner that was both rapamycin-sensitive (proteasome activity; phosphorylation of S6K1 and rpS6) and rapamycin-insensitive (phosphorylation of eEF2, ERK1/2 and UBF; gene expression of the myostatin target Mighty as well as c-Myc and its targets involved in ribosome biogenesis). The role of c-Myc was tested in vitro using the inhibitor 10058-F4, which, over time, decreased basal RNA and MPS in a dose-dependent manner (correlation of RNA and MPS, r(2) = 0.98), even though it had no effect on the acute stimulation of protein synthesis. In conclusion, acute resistance exercise stimulated rapamycin-sensitive and -insensitive mechanisms that regulate translation activity and capacity.
© 2015 The Authors. The Journal of Physiology © 2015 The Physiological Society.
Figures
References
-
- Adams GR, Caiozzo VJ, Haddad F & Baldwin KM (2002). Cellular and molecular responses to increased skeletal muscle loading after irradiation. Am J Physiol Cell Physiol 283, C1182–1195. - PubMed
-
- Akiho M, Nakashima H, Sakata M, Yamasa Y, Yamaguchi A & Sakuma K (2010). Expression profile of Notch‐1 in mechanically overloaded plantaris muscle of mice. Life Sci 86, 59–65. - PubMed
-
- Al‐Khalili L, de Castro Barbosa T, Ostling J, Massart J, Cuesta PG, Osler ME, Katayama M, Nystrom AC, Oscarsson J & Zierath JR (2014). Proteasome inhibition in skeletal muscle cells unmasks metabolic derangements in type 2 diabetes. Am J Physiol Cell Physiol 307, C774–787. - PubMed
-
- Arabi A, Wu S, Ridderstrale K, Bierhoff H, Shiue C, Fatyol K, Fahlen S, Hydbring P, Soderberg O, Grummt I, Larsson LG & Wright AP (2005). c‐Myc associates with ribosomal DNA and activates RNA polymerase I transcription. Nat Cell Biol 7, 303–310. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous