

Piezo2 is the principal mechanotransduction channel for proprioception
- PMID: 26551544
- PMCID: PMC4661126
- DOI: 10.1038/nn.4162
Piezo2 is the principal mechanotransduction channel for proprioception
Abstract
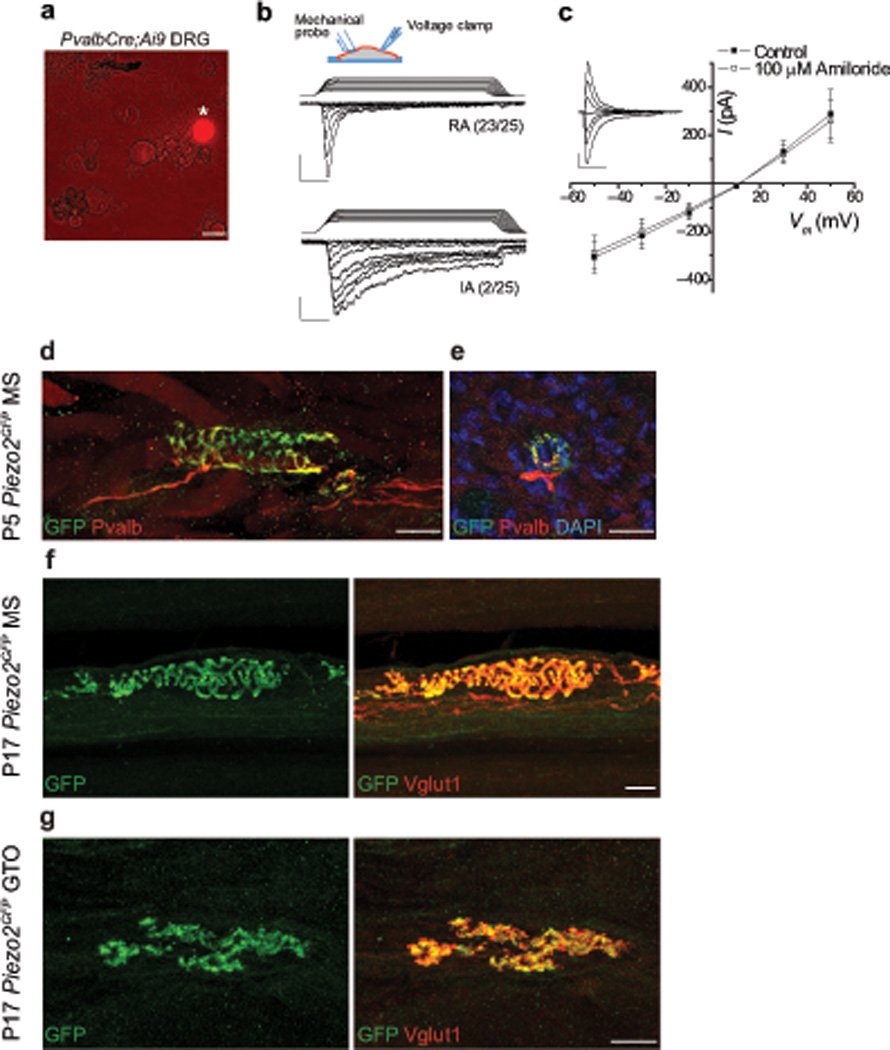
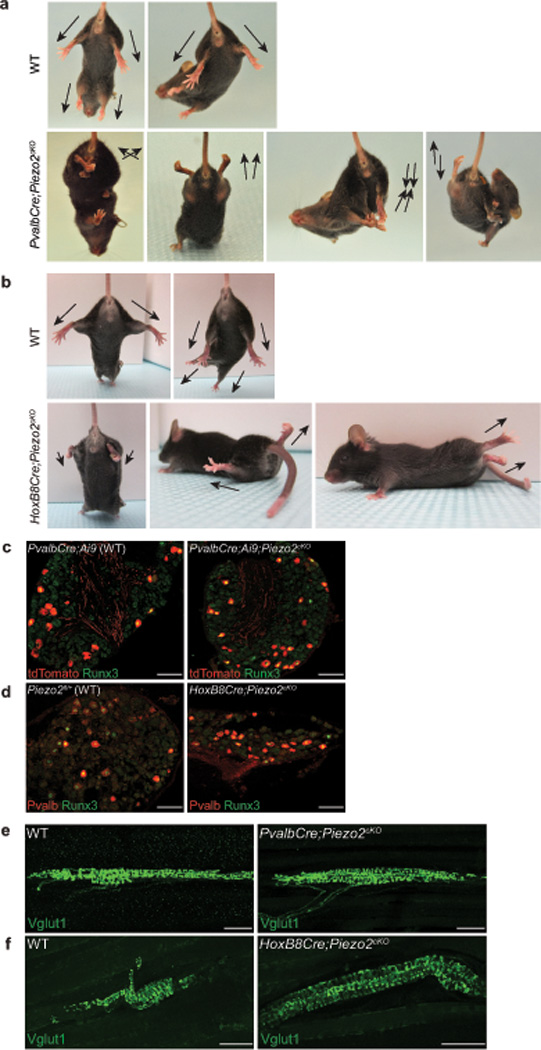
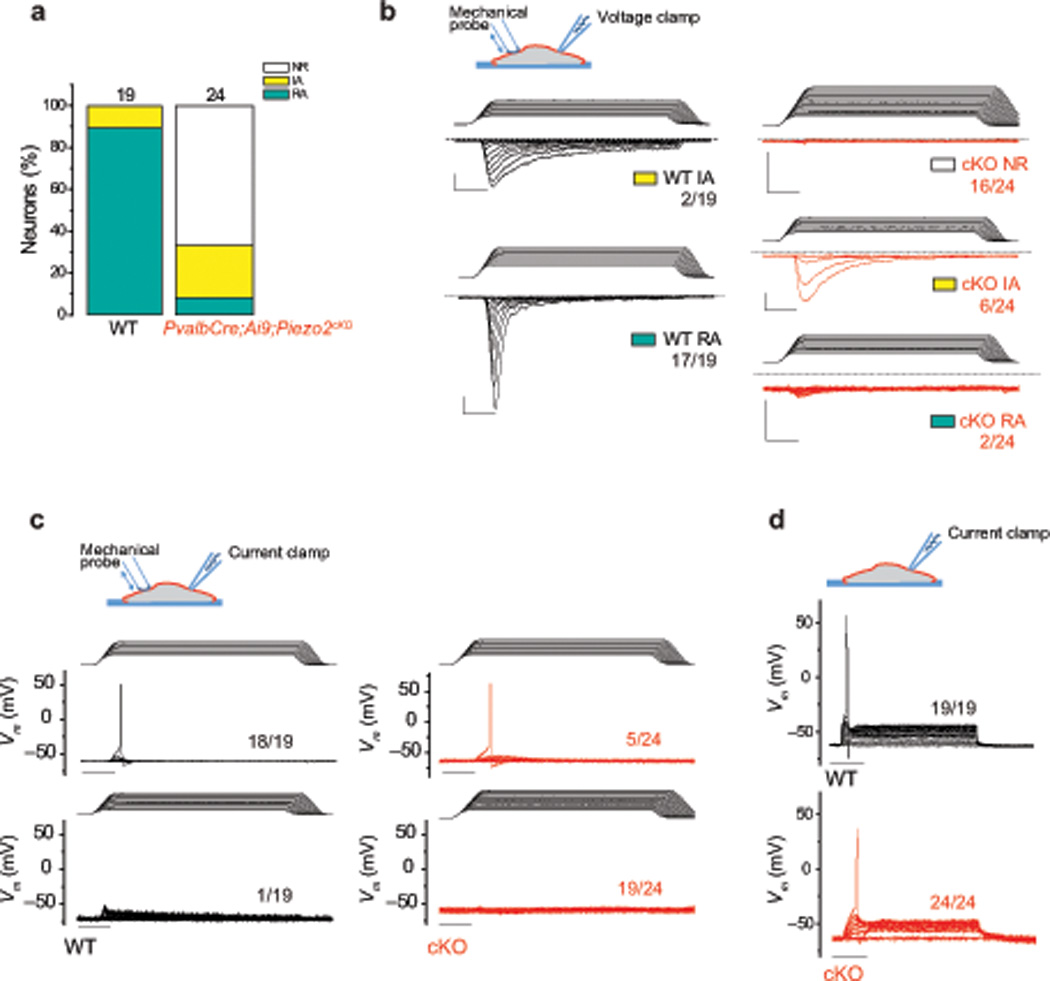
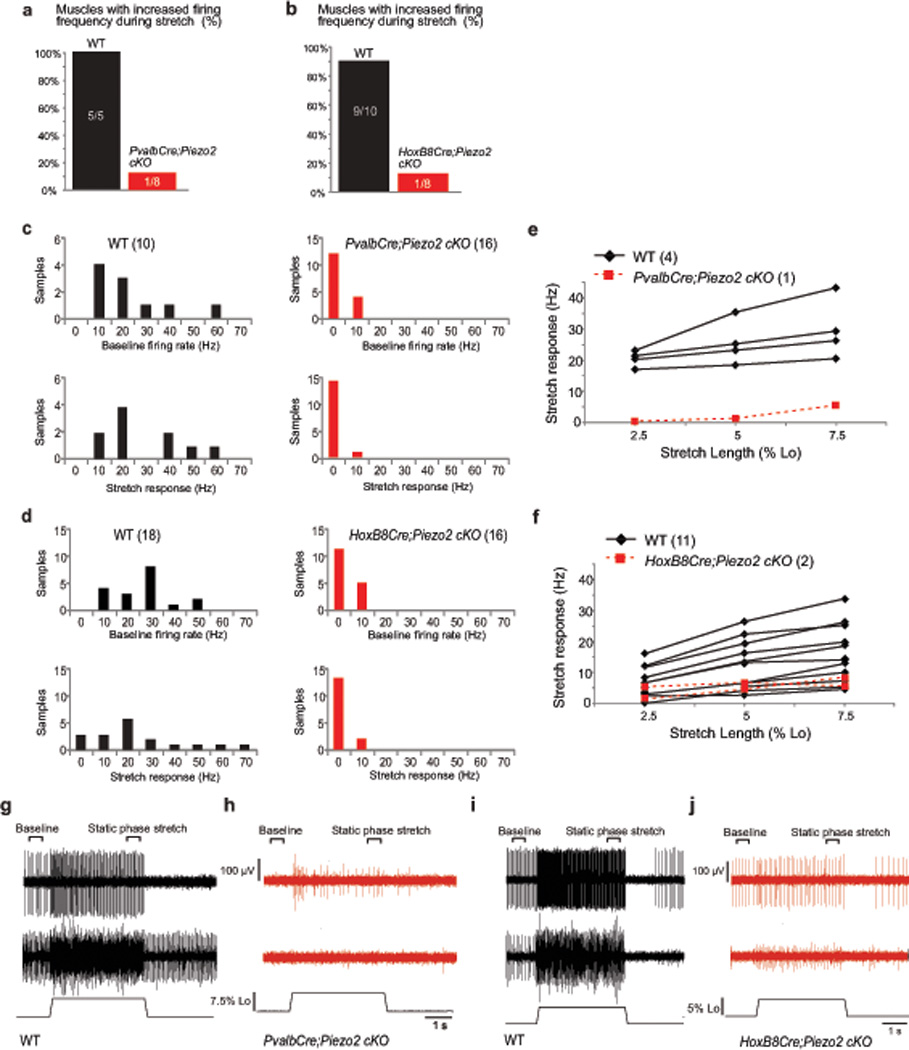
Proprioception, the perception of body and limb position, is mediated by proprioceptors, specialized mechanosensory neurons that convey information about the stretch and tension experienced by muscles, tendons, skin and joints. In mammals, the molecular identity of the stretch-sensitive channel that mediates proprioception is unknown. We found that the mechanically activated nonselective cation channel Piezo2 was expressed in sensory endings of proprioceptors innervating muscle spindles and Golgi tendon organs in mice. Two independent mouse lines that lack Piezo2 in proprioceptive neurons showed severely uncoordinated body movements and abnormal limb positions. Moreover, the mechanosensitivity of parvalbumin-expressing neurons that predominantly mark proprioceptors was dependent on Piezo2 expression in vitro, and the stretch-induced firing of proprioceptors in muscle-nerve recordings was markedly reduced in Piezo2-deficient mice. Together, our results indicate that Piezo2 is the major mechanotransducer of mammalian proprioceptors.
Figures
References
-
- Sherrington C. On the proprio-ceptive system, especially in its reflex aspect. Brain. 1907;29:467–482.
-
- Proske U, Gandevia SC. The proprioceptive senses: their roles in signaling body shape, body position and movement, and muscle force. Physiological reviews. 2012;92:1651–1697. - PubMed
-
- Walker RG, Willingham AT, Zuker CSA. Drosophila mechanosensory transduction channel. Science. 2000;287:2229–2234. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
