

Molecular systems evaluation of oligomerogenic APP(E693Q) and fibrillogenic APP(KM670/671NL)/PSEN1(Δexon9) mouse models identifies shared features with human Alzheimer's brain molecular pathology
- PMID: 26552589
- PMCID: PMC4862938
- DOI: 10.1038/mp.2015.167
Molecular systems evaluation of oligomerogenic APP(E693Q) and fibrillogenic APP(KM670/671NL)/PSEN1(Δexon9) mouse models identifies shared features with human Alzheimer's brain molecular pathology
Erratum in
-
Molecular systems evaluation of oligomerogenic APP(E693Q) and fibrillogenic APP(KM670/671NL)/PSEN1(Δexon9) mouse models identifies shared features with human Alzheimer's brain molecular pathology.Mol Psychiatry. 2016 Aug;21(8):1153-4. doi: 10.1038/mp.2015.215. Epub 2016 Jan 19. Mol Psychiatry. 2016. PMID: 26782051 Free PMC article. No abstract available.
Abstract
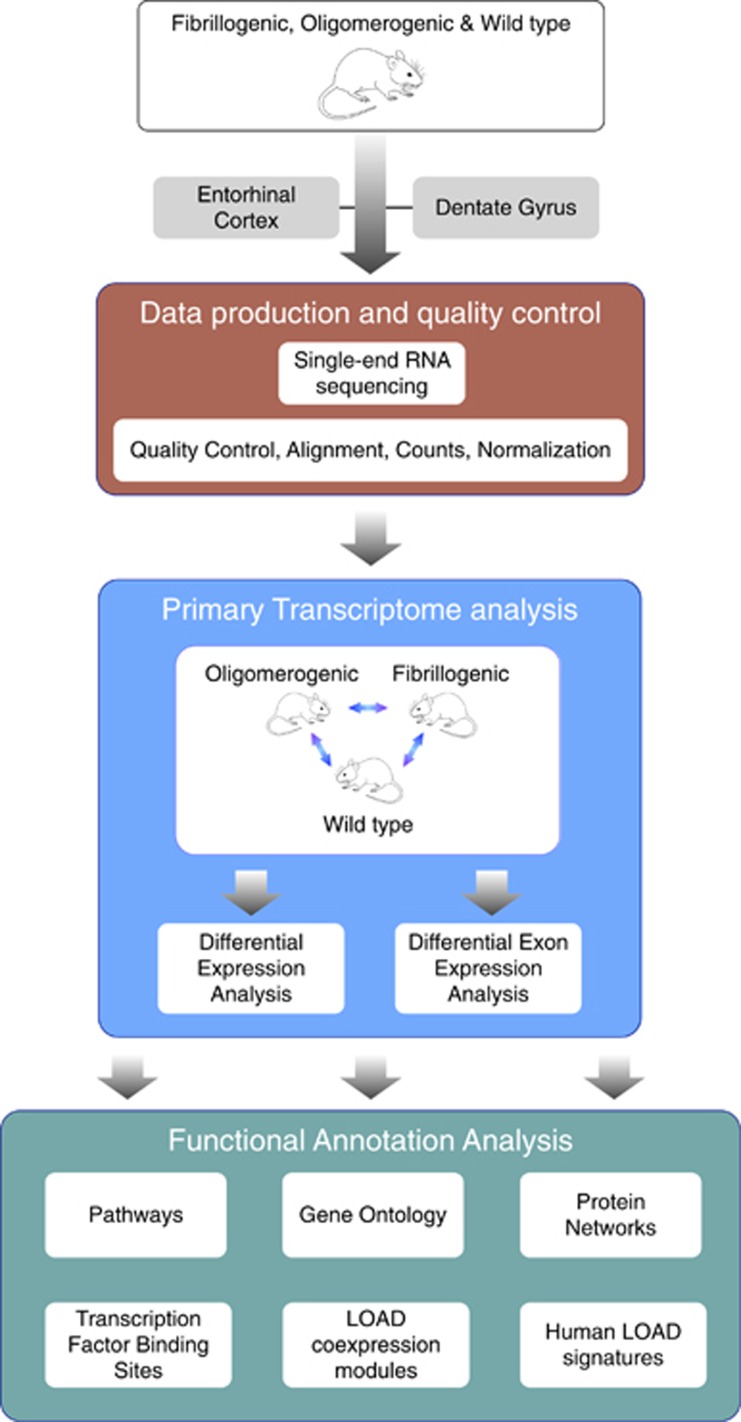
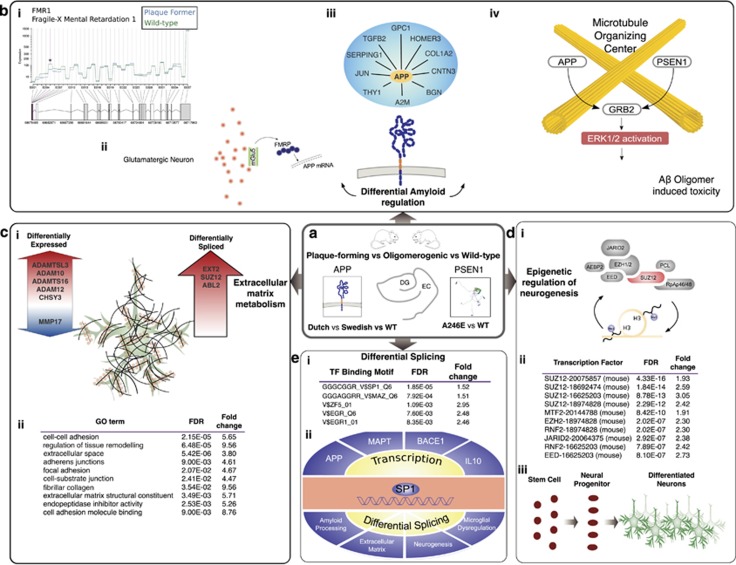
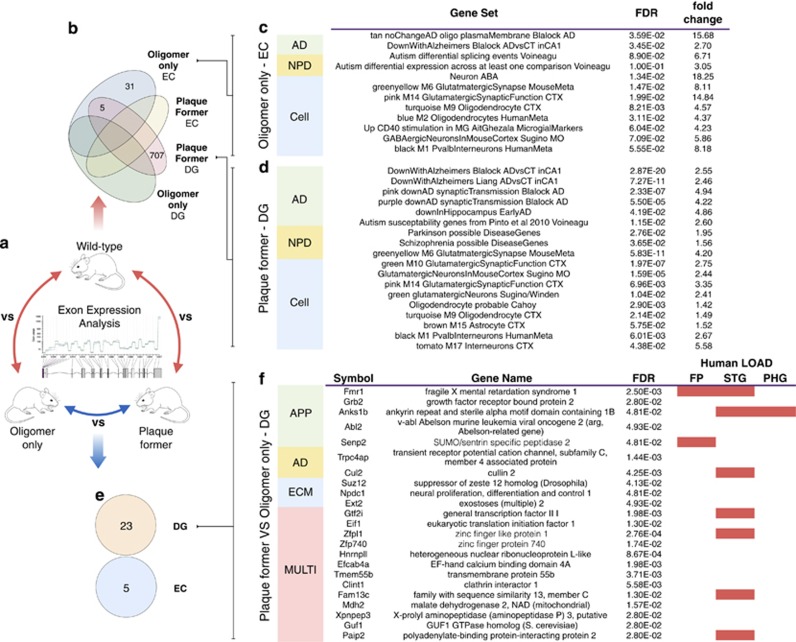
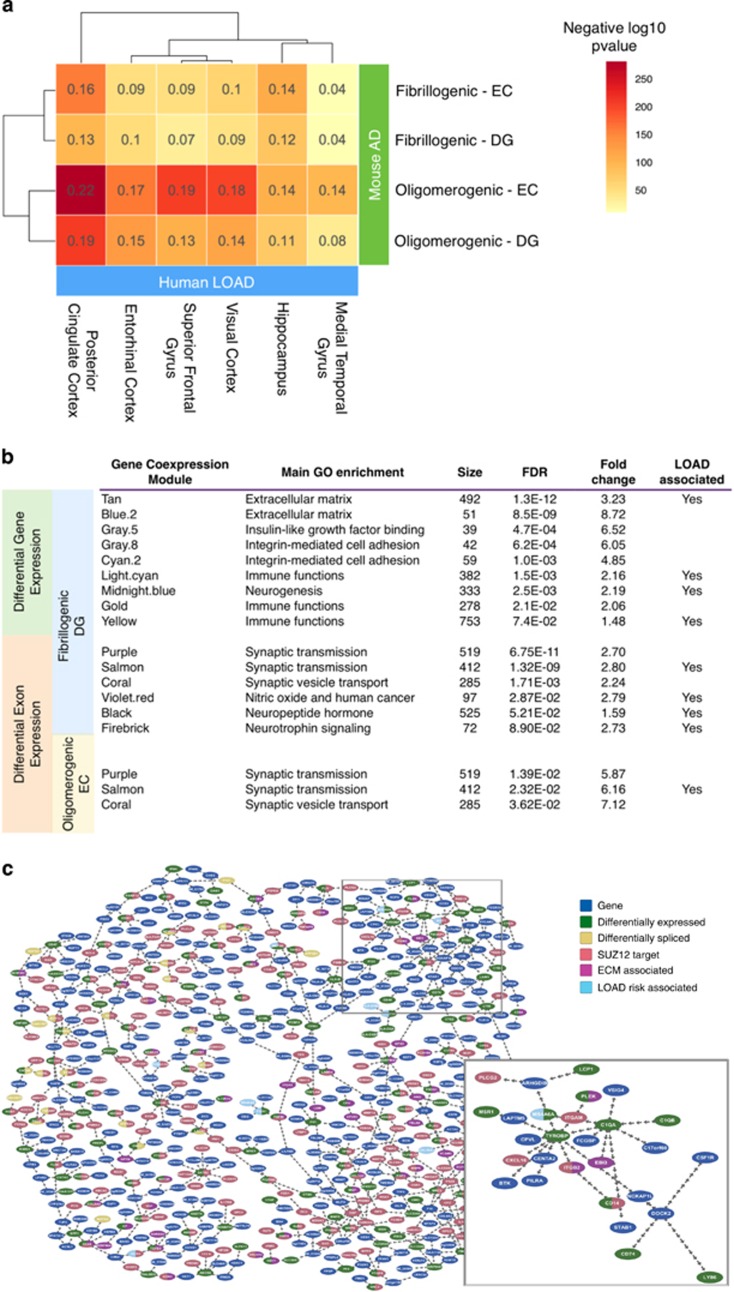
Identification and characterization of molecular mechanisms that connect genetic risk factors to initiation and evolution of disease pathophysiology represent major goals and opportunities for improving therapeutic and diagnostic outcomes in Alzheimer's disease (AD). Integrative genomic analysis of the human AD brain transcriptome holds potential for revealing novel mechanisms of dysfunction that underlie the onset and/or progression of the disease. We performed an integrative genomic analysis of brain tissue-derived transcriptomes measured from two lines of mice expressing distinct mutant AD-related proteins. The first line expresses oligomerogenic mutant APP(E693Q) inside neurons, leading to the accumulation of amyloid beta (Aβ) oligomers and behavioral impairment, but never develops parenchymal fibrillar amyloid deposits. The second line expresses APP(KM670/671NL)/PSEN1(Δexon9) in neurons and accumulates fibrillar Aβ amyloid and amyloid plaques accompanied by neuritic dystrophy and behavioral impairment. We performed RNA sequencing analyses of the dentate gyrus and entorhinal cortex from each line and from wild-type mice. We then performed an integrative genomic analysis to identify dysregulated molecules and pathways, comparing transgenic mice with wild-type controls as well as to each other. We also compared these results with datasets derived from human AD brain. Differential gene and exon expression analysis revealed pervasive alterations in APP/Aβ metabolism, epigenetic control of neurogenesis, cytoskeletal organization and extracellular matrix (ECM) regulation. Comparative molecular analysis converged on FMR1 (Fragile X Mental Retardation 1), an important negative regulator of APP translation and oligomerogenesis in the post-synaptic space. Integration of these transcriptomic results with human postmortem AD gene networks, differential expression and differential splicing signatures identified significant similarities in pathway dysregulation, including ECM regulation and neurogenesis, as well as strong overlap with AD-associated co-expression network structures. The strong overlap in molecular systems features supports the relevance of these findings from the AD mouse models to human AD.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
