

Multiple Roles for Nogo Receptor 1 in Visual System Plasticity
- PMID: 26552866
- PMCID: PMC6592612
- DOI: 10.1177/1073858415614564
Multiple Roles for Nogo Receptor 1 in Visual System Plasticity
Abstract
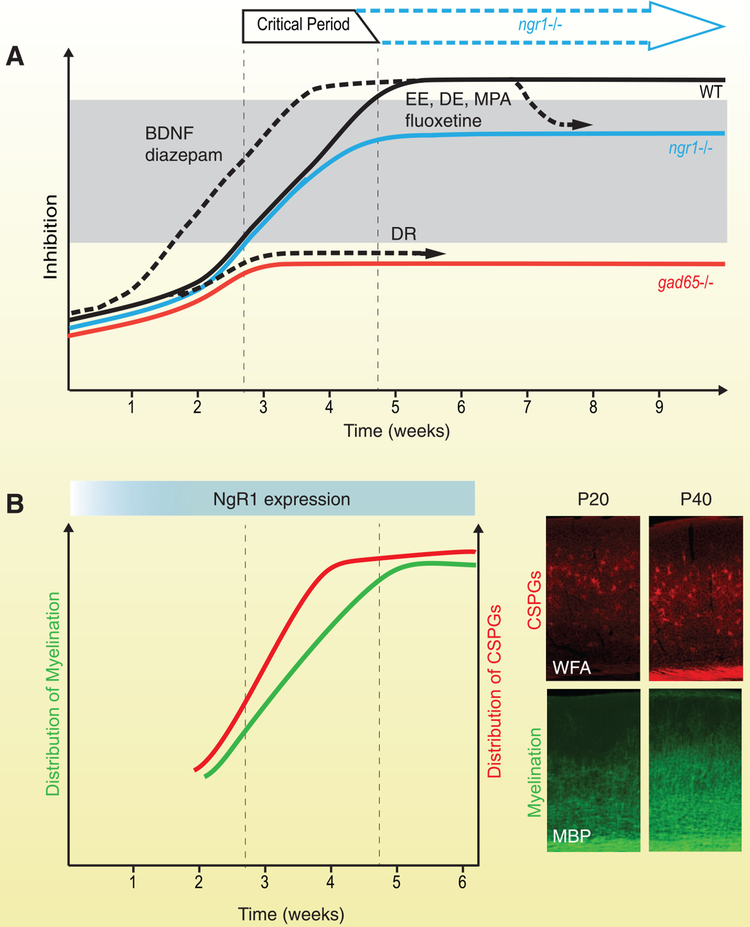
During the developmental critical period for visual plasticity, discordant vision alters the responsiveness of neurons in visual cortex. The subsequent closure of the critical period not only consolidates neural function but also limits recovery of acuity from preceding abnormal visual experience. Despite species-specific differences in circuitry of the visual system, these characteristics are conserved. The nogo-66 receptor 1 (ngr1) is one of only a small number of genes identified thus far that is essential to closing the critical period. Mice lacking a functional ngr1 gene retain developmental visual plasticity as adults and their visual acuity spontaneously improves after prolonged visual deprivation. Experiments employing conditional mouse genetics have revealed that ngr1 restricts plasticity within distinct circuits for ocular dominance and visual acuity. However, the mechanisms by which NgR1 limits plasticity have not been elucidated, in part because the subcellular localization and signal transduction of the protein are only partially understood. Here we explore potential mechanisms for NgR1 function in relation to manipulations that reactivate visual plasticity in adults and propose lines of investigation to address relevant gaps in knowledge.
Keywords: amblyopia; critical period; dendritic spines; mouse; myelin; ocular dominance; visual acuity; visual cortex; visual plasticity.
© The Author(s) 2015.
Conflict of interest statement
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Figures
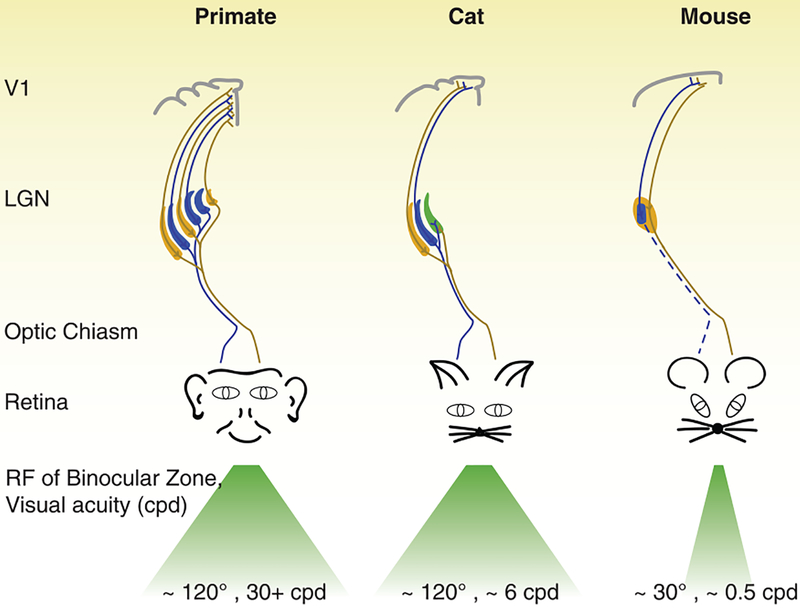
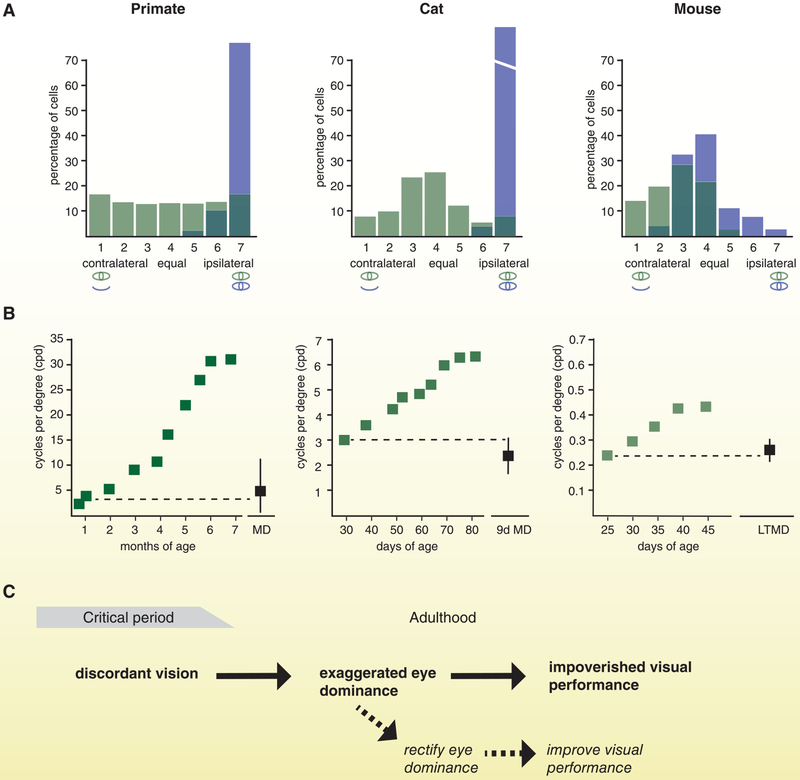
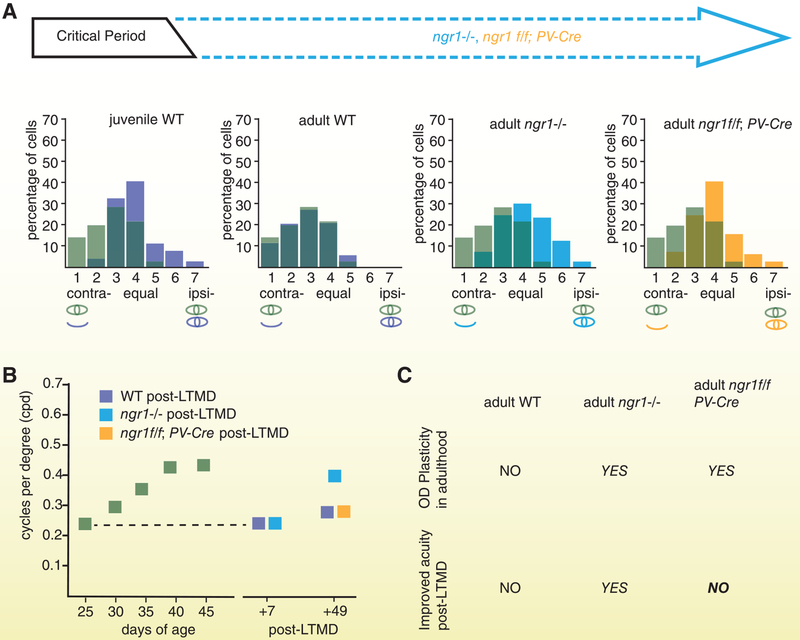
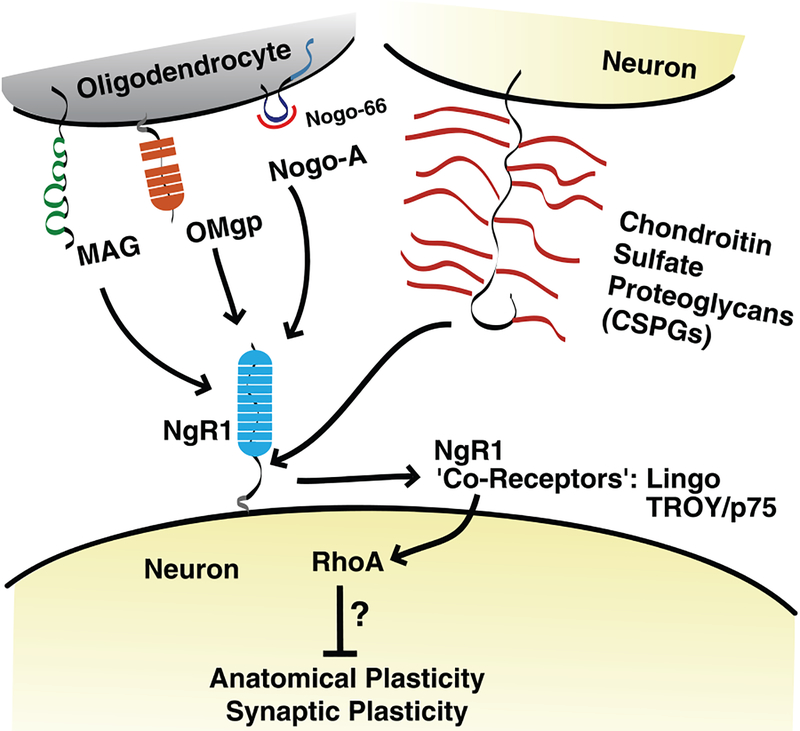

References
-
- Antonini A, Stryker MP. 1993. Rapid remodeling of axonal arbors in the visual cortex. Science 260:1819–21. - PubMed
-
- Baroncelli L, Sale A, Viegi A, Maya Vetencourt JF, De Pasquale R, Baldini S, and others. 2010. Experience-dependent reactivation of ocular dominance plasticity in the adult visual cortex. Exp Neurol 226:100–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
