

Pannexin-1 and P2X7-Receptor Are Required for Apoptotic Osteocytes in Fatigued Bone to Trigger RANKL Production in Neighboring Bystander Osteocytes
- PMID: 26553756
- PMCID: PMC4915221
- DOI: 10.1002/jbmr.2740
Pannexin-1 and P2X7-Receptor Are Required for Apoptotic Osteocytes in Fatigued Bone to Trigger RANKL Production in Neighboring Bystander Osteocytes
Abstract
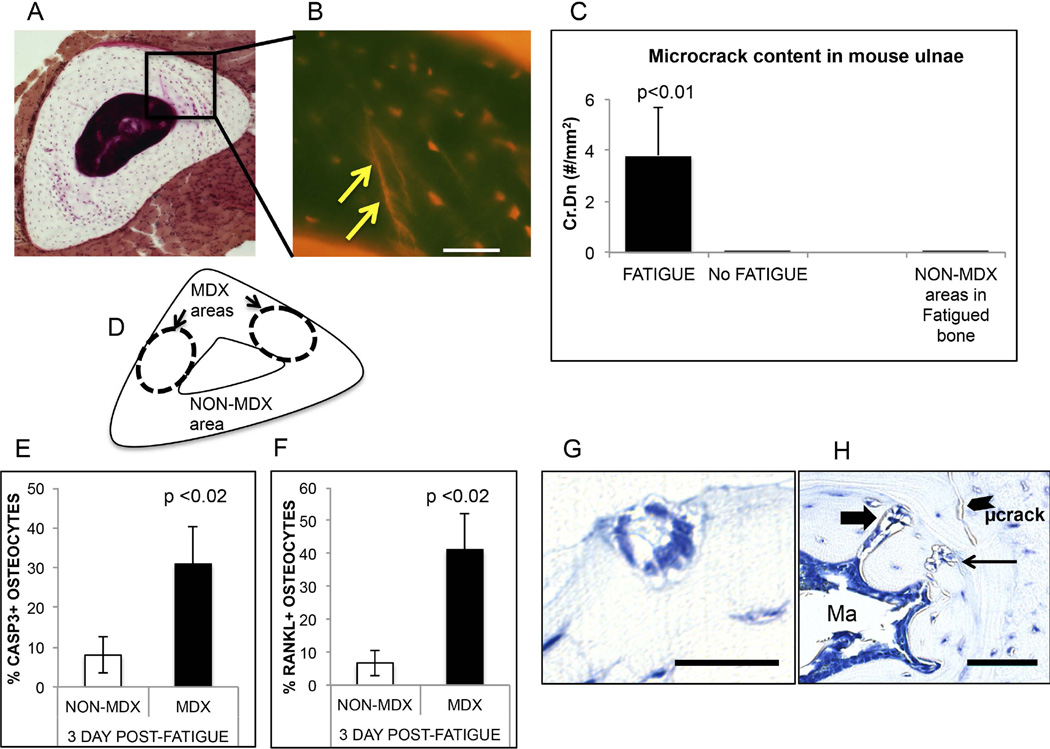
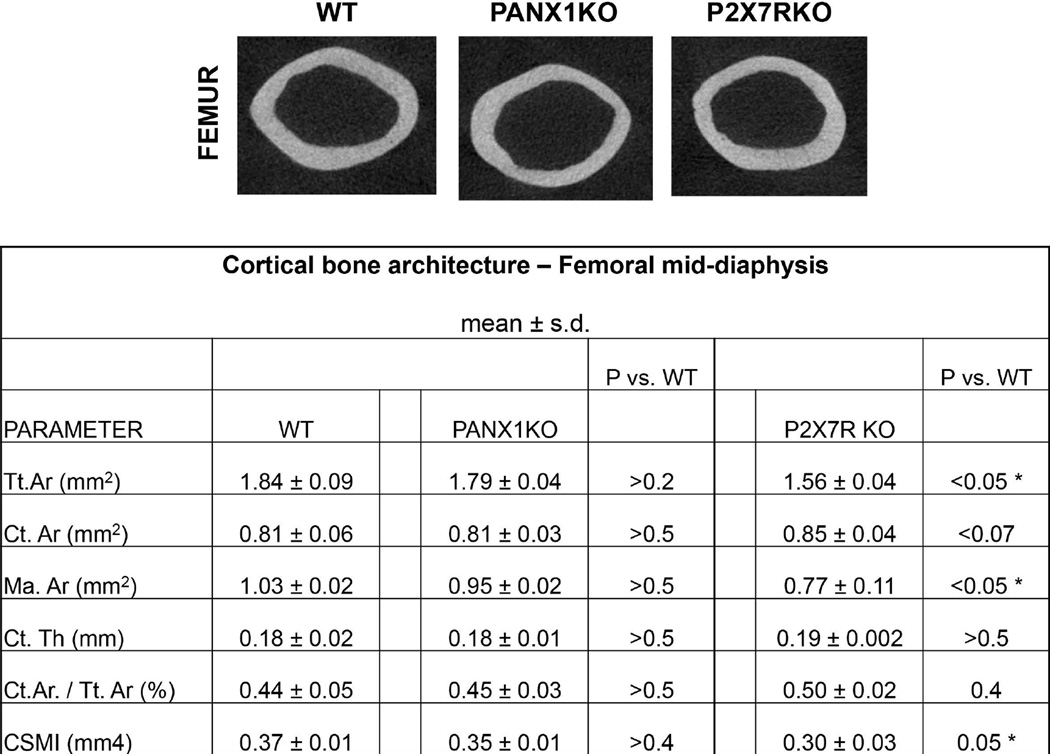
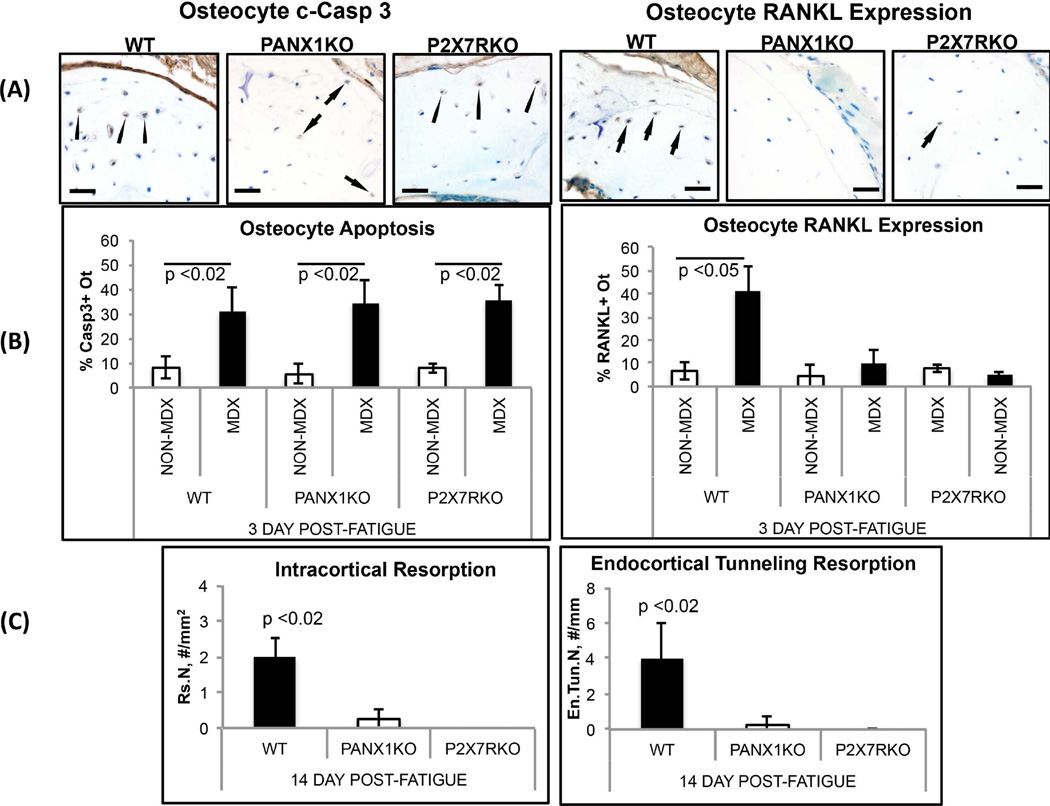
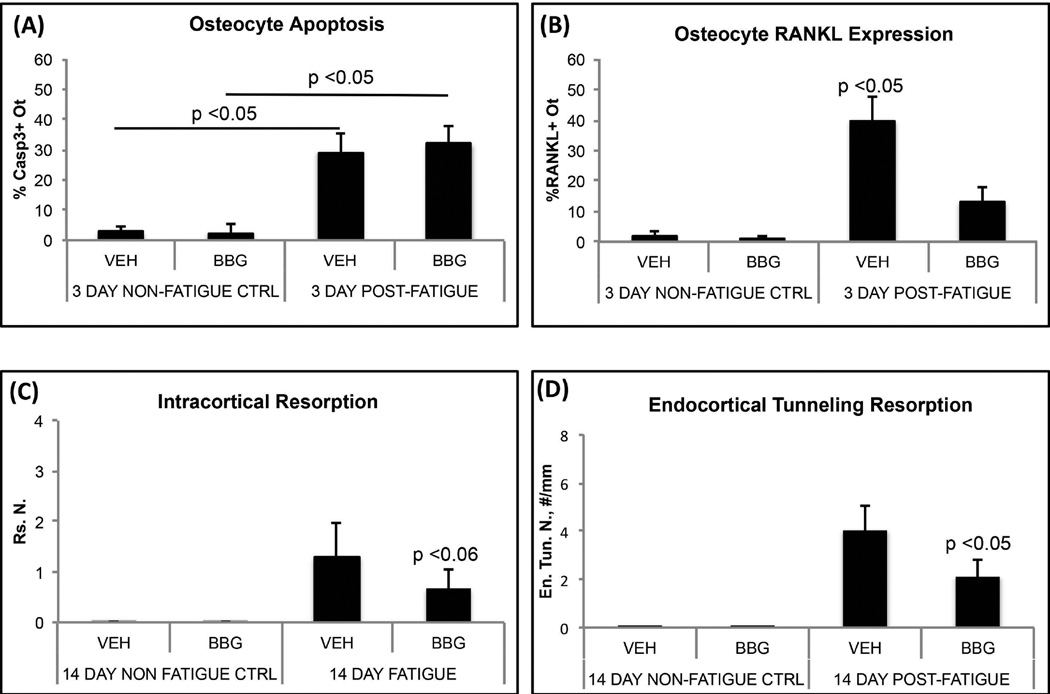
Osteocyte apoptosis is required to induce intracortical bone remodeling after microdamage in animal models, but how apoptotic osteocytes signal neighboring "bystander" cells to initiate the remodeling process is unknown. Apoptosis has been shown to open pannexin-1 (Panx1) channels to release adenosine diphosphate (ATP) as a "find-me" signal for phagocytic cells. To address whether apoptotic osteocytes use this signaling mechanism, we adapted the rat ulnar fatigue-loading model to reproducibly introduce microdamage into mouse cortical bone and measured subsequent changes in osteocyte apoptosis, receptor activator of NF-κB ligand (RANKL) expression and osteoclastic bone resorption in wild-type (WT; C57Bl/6) mice and in mice genetically deficient in Panx1 (Panx1KO). Mouse ulnar loading produced linear microcracks comparable in number and location to the rat model. WT mice showed increased osteocyte apoptosis and RANKL expression at microdamage sites at 3 days after loading and increased intracortical remodeling and endocortical tunneling at day 14. With fatigue, Panx1KO mice exhibited levels of microdamage and osteocyte apoptosis identical to WT mice. However, they did not upregulate RANKL in bystander osteocytes or initiate resorption. Panx1 interacts with P2X7 R in ATP release; thus, we examined P2X7 R-deficient mice and WT mice treated with P2X7 R antagonist Brilliant Blue G (BBG) to test the possible role of ATP as a find-me signal. P2X7 RKO mice failed to upregulate RANKL in osteocytes or induce resorption despite normally elevated osteocyte apoptosis after fatigue loading. Similarly, treatment of fatigued C57Bl/6 mice with BBG mimicked behavior of both Panx1KO and P2X7 RKO mice; BBG had no effect on osteocyte apoptosis in fatigued bone but completely prevented increases in bystander osteocyte RANKL expression and attenuated activation of resorption by more than 50%. These results indicate that activation of Panx1 and P2X7 R are required for apoptotic osteocytes in fatigued bone to trigger RANKL production in neighboring bystander osteocytes and implicate ATP as an essential signal mediating this process.
Keywords: BYSTANDER SIGNALING; FATIGUE MICRODAMAGE; OSTEOCYTE APOPTOSIS; P2X7 RECEPTOR; PANNEXIN 1; RANKL.
© 2016 American Society for Bone and Mineral Research.
Figures
Similar articles
-
Apoptotic Osteocytes Induce RANKL Production in Bystanders via Purinergic Signaling and Activation of Pannexin Channels.J Bone Miner Res. 2020 May;35(5):966-977. doi: 10.1002/jbmr.3954. Epub 2020 Feb 11. J Bone Miner Res. 2020. PMID: 31910292 Free PMC article.
-
Osteocyte Apoptosis Caused by Hindlimb Unloading is Required to Trigger Osteocyte RANKL Production and Subsequent Resorption of Cortical and Trabecular Bone in Mice Femurs.J Bone Miner Res. 2016 Jul;31(7):1356-65. doi: 10.1002/jbmr.2807. Epub 2016 Mar 22. J Bone Miner Res. 2016. PMID: 26852281 Free PMC article.
-
Activation of resorption in fatigue-loaded bone involves both apoptosis and active pro-osteoclastogenic signaling by distinct osteocyte populations.Bone. 2012 May;50(5):1115-22. doi: 10.1016/j.bone.2012.01.025. Epub 2012 Feb 9. Bone. 2012. PMID: 22342796 Free PMC article.
-
Regulatory mechanisms of RANKL presentation to osteoclast precursors.Curr Osteoporos Rep. 2014 Mar;12(1):115-20. doi: 10.1007/s11914-014-0189-0. Curr Osteoporos Rep. 2014. PMID: 24477414 Review.
-
Pannexin-1 channels in epilepsy.Neurosci Lett. 2019 Mar 16;695:71-75. doi: 10.1016/j.neulet.2017.09.004. Epub 2017 Sep 5. Neurosci Lett. 2019. PMID: 28886985 Review.
Cited by
-
Strategies of Macrophages to Maintain Bone Homeostasis and Promote Bone Repair: A Narrative Review.J Funct Biomater. 2022 Dec 29;14(1):18. doi: 10.3390/jfb14010018. J Funct Biomater. 2022. PMID: 36662065 Free PMC article. Review.
-
Role of pannexin 1 channels in load-induced skeletal response.Ann N Y Acad Sci. 2019 Apr;1442(1):79-90. doi: 10.1111/nyas.13914. Epub 2018 Jun 28. Ann N Y Acad Sci. 2019. PMID: 29952014 Free PMC article.
-
Connexins and Pannexins in Bone and Skeletal Muscle.Curr Osteoporos Rep. 2017 Aug;15(4):326-334. doi: 10.1007/s11914-017-0374-z. Curr Osteoporos Rep. 2017. PMID: 28647887 Free PMC article. Review.
-
Is There a Governing Role of Osteocytes in Bone Tissue Regeneration?Curr Osteoporos Rep. 2020 Oct;18(5):541-550. doi: 10.1007/s11914-020-00610-6. Curr Osteoporos Rep. 2020. PMID: 32676786 Free PMC article. Review.
-
The osteocyte and its osteoclastogenic potential.Front Endocrinol (Lausanne). 2023 May 24;14:1121727. doi: 10.3389/fendo.2023.1121727. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 37293482 Free PMC article. Review.
References
-
- Aguirre J, Plotkin LI, Stewart SA, Weinstein RS, Parfitt AM, Manolagas SC, Bellido T. Osteocyte apoptosis is induced by weightlessness in mice and precedes osteoclast recruitment and bone loss. Journal of bone and mineral research. 2006;21(4):605–615. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
