

Exosomes from HIV-1-infected Cells Stimulate Production of Pro-inflammatory Cytokines through Trans-activating Response (TAR) RNA
- PMID: 26553869
- PMCID: PMC4714213
- DOI: 10.1074/jbc.M115.662171
Exosomes from HIV-1-infected Cells Stimulate Production of Pro-inflammatory Cytokines through Trans-activating Response (TAR) RNA
Abstract
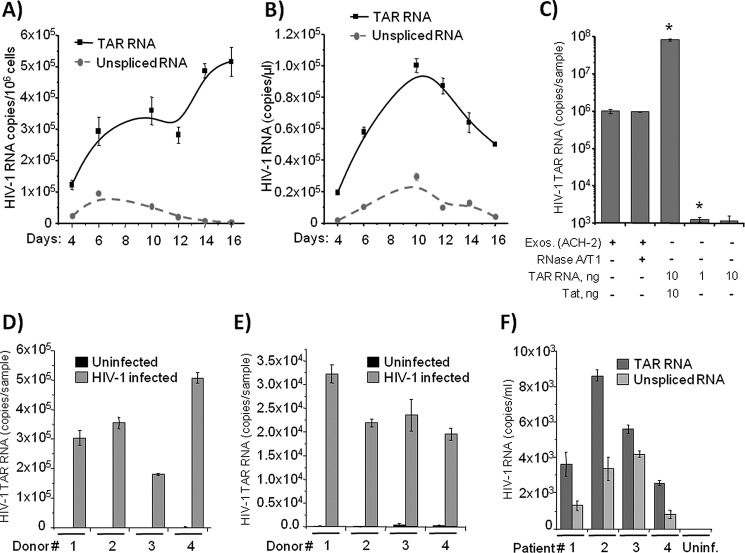
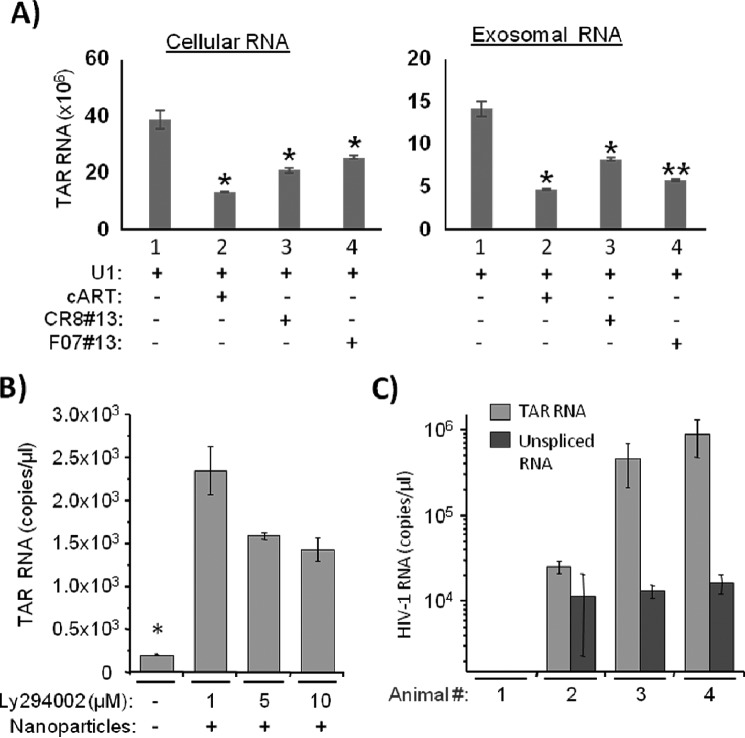
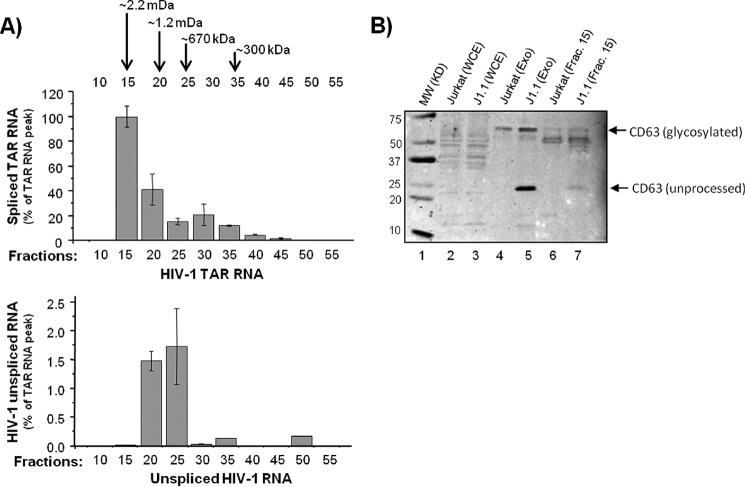
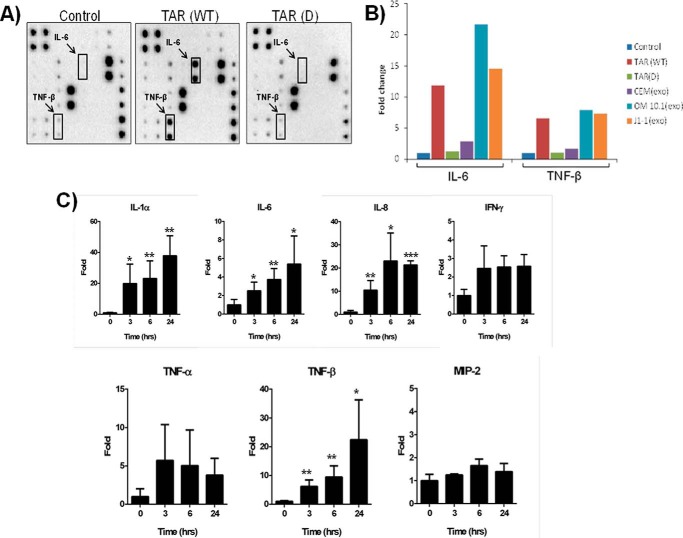
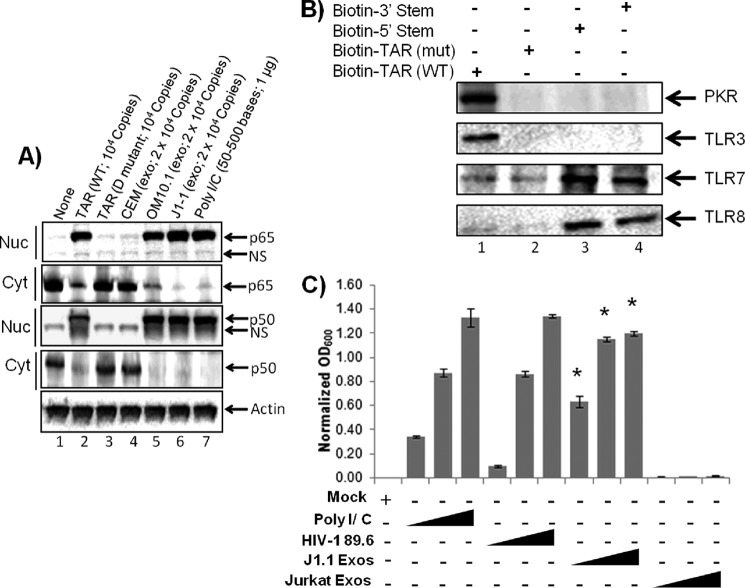
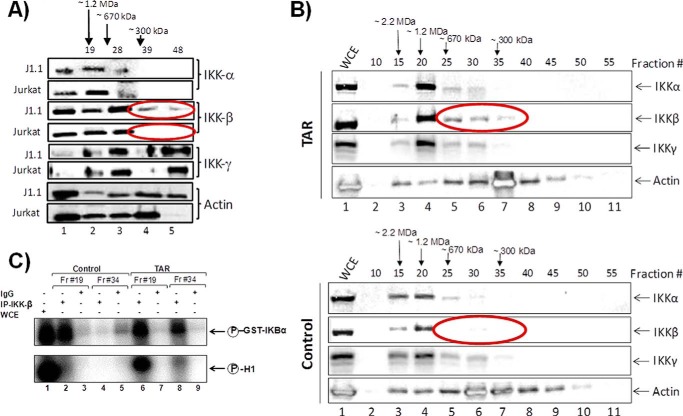
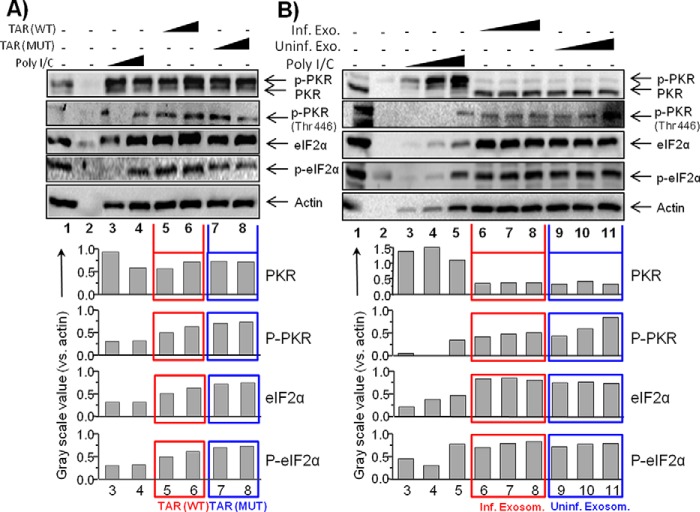
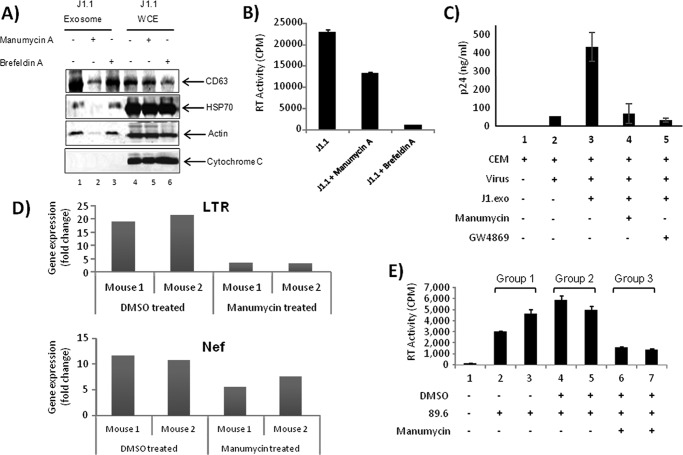
HIV-1 infection results in a chronic illness because long-term highly active antiretroviral therapy can lower viral titers to an undetectable level. However, discontinuation of therapy rapidly increases virus burden. Moreover, patients under highly active antiretroviral therapy frequently develop various metabolic disorders, neurocognitive abnormalities, and cardiovascular diseases. We have previously shown that exosomes containing trans-activating response (TAR) element RNA enhance susceptibility of undifferentiated naive cells to HIV-1 infection. This study indicates that exosomes from HIV-1-infected primary cells are highly abundant with TAR RNA as detected by RT-real time PCR. Interestingly, up to a million copies of TAR RNA/μl were also detected in the serum from HIV-1-infected humanized mice suggesting that TAR RNA may be stable in vivo. Incubation of exosomes from HIV-1-infected cells with primary macrophages resulted in a dramatic increase of proinflammatory cytokines, IL-6 and TNF-β, indicating that exosomes containing TAR RNA could play a direct role in control of cytokine gene expression. The intact TAR molecule was able to bind to PKR and TLR3 effectively, whereas the 5' and 3' stems (TAR microRNAs) bound best to TLR7 and -8 and none to PKR. Binding of TAR to PKR did not result in its phosphorylation, and therefore, TAR may be a dominant negative decoy molecule in cells. The TLR binding through either TAR RNA or TAR microRNA potentially can activate the NF-κB pathway and regulate cytokine expression. Collectively, these results imply that exosomes containing TAR RNA could directly affect the proinflammatory cytokine gene expression and may explain a possible mechanism of inflammation observed in HIV-1-infected patients under cART.
Keywords: TAR RNA; cytokinesis; exosome (vesicle); human immunodeficiency virus (HIV); macrophage; protein kinase RNA-activated (PKR); toll-like receptor (TLR).
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- UNAIDS (2012) UNAIDS Report on the Global AIDS Epidemic 2012. http://www.unaids.org/sites/default/files/media_asset/20121120_UNAIDS_Gl...
-
- van der Sluis R. M., Jeeninga R. E., and Berkhout B. (2013) Establishment and molecular mechanisms of HIV-1 latency in T cells. Curr. Opin. Virol. 3, 700–706 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
