

The sequence of sequencers: The history of sequencing DNA
- PMID: 26554401
- PMCID: PMC4727787
- DOI: 10.1016/j.ygeno.2015.11.003
The sequence of sequencers: The history of sequencing DNA
Abstract
Determining the order of nucleic acid residues in biological samples is an integral component of a wide variety of research applications. Over the last fifty years large numbers of researchers have applied themselves to the production of techniques and technologies to facilitate this feat, sequencing DNA and RNA molecules. This time-scale has witnessed tremendous changes, moving from sequencing short oligonucleotides to millions of bases, from struggling towards the deduction of the coding sequence of a single gene to rapid and widely available whole genome sequencing. This article traverses those years, iterating through the different generations of sequencing technology, highlighting some of the key discoveries, researchers, and sequences along the way.
Keywords: DNA; History; RNA; Sequencer; Sequencing.
Copyright © 2015 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
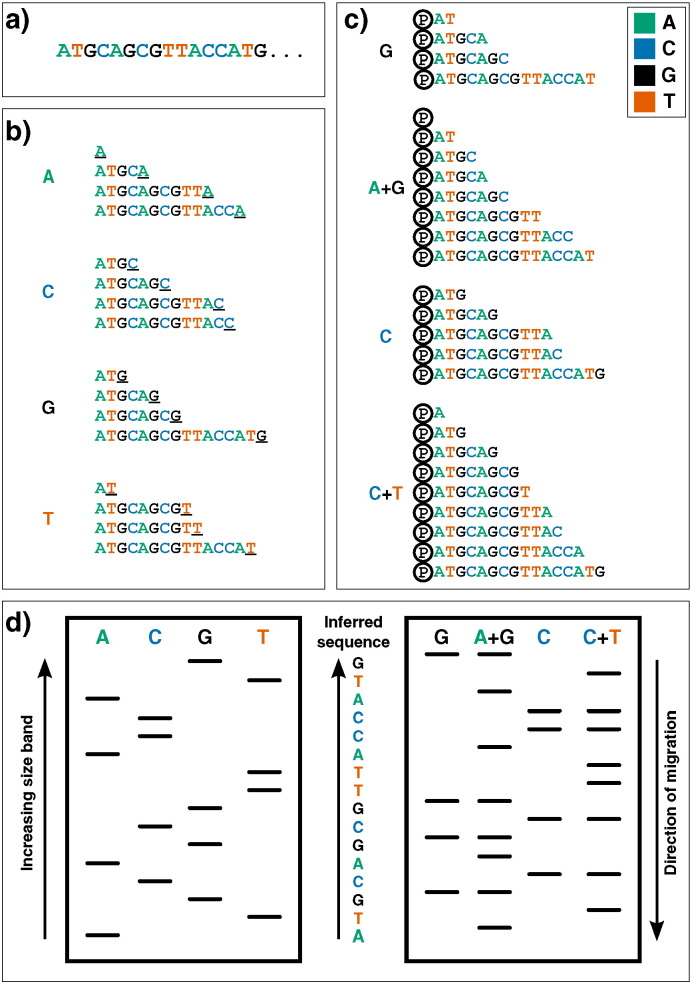
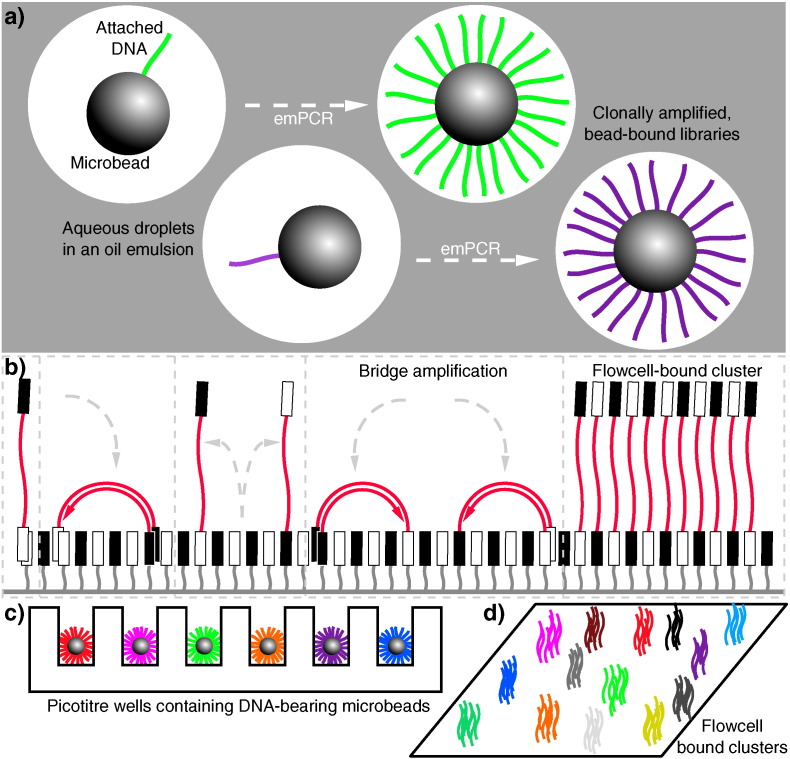
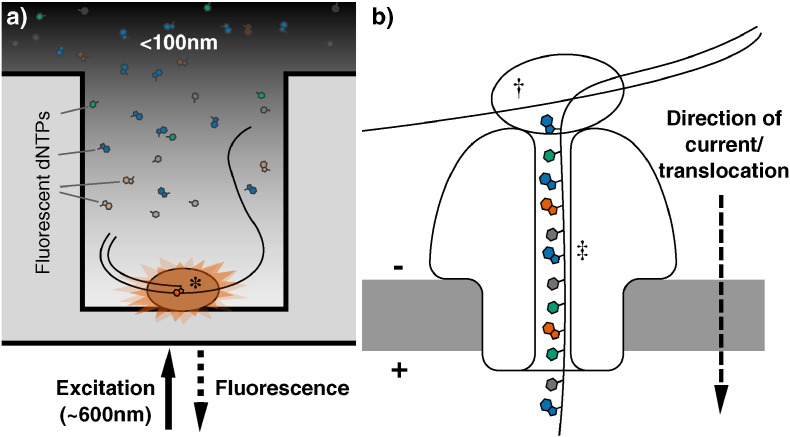
References
-
- Sanger F. 1980. Frederick Sanger — Biographical. (URL http://www.nobelprize.org/nobel_prizes/chemistry/laureates/1980/sanger-b...)
-
- Watson J., Crick F. Molecular structure of nucleic acids. Nature. 1953;171:709–756. (URL http://www.nature.com/physics/looking-back/crick/) - PubMed
-
- Zallen D.T. Despite Franklin's work, Wilkins earned his Nobel. Nature. 2003;425:15. - PubMed
-
- Holley R.W., Apgar J., Merrill S.H., Zubkoff P.L. Nucleotide and oligonucleotide compositions of the alanine-, valine-, and tyrosine-acceptor soluble ribonucleic acids of yeast. J. Am. Chem. Soc. 1961;83:4861–4862. (URL http://pubs.acs.org/doi/abs/10.1021/ja01484a040) - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
