

An Engineered Minimal WASP-Myosin Fusion Protein Reveals Essential Functions for Endocytosis
- PMID: 26555049
- PMCID: PMC4643393
- DOI: 10.1016/j.devcel.2015.10.007
An Engineered Minimal WASP-Myosin Fusion Protein Reveals Essential Functions for Endocytosis
Abstract
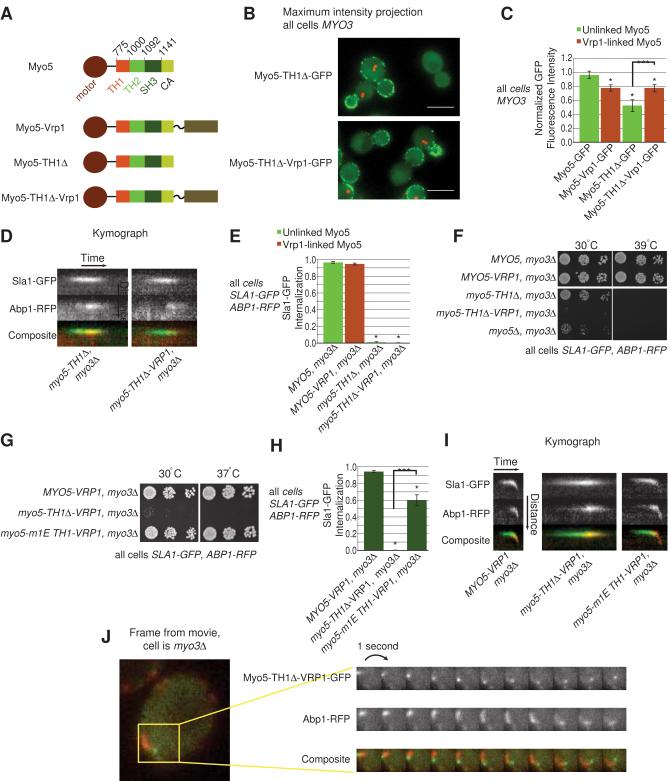
Actin polymerization powers membrane deformation during many processes, including clathrin-mediated endocytosis (CME). During CME in yeast, actin polymerization is triggered and coordinated by a six-protein WASP/Myosin complex that includes WASP, class I myosins (Myo3 and Myo5), WIP (Vrp1), and two other proteins. We show that a single engineered protein can replace this entire complex while still supporting CME. This engineered protein reveals that the WASP/Myosin complex has four essential activities: recruitment to endocytic sites, anchorage to the plasma membrane, Arp2/3 activation, and transient actin filament binding by the motor domain. The requirement for both membrane and F-actin binding reveals that myosin-mediated coupling between actin filaments and the base of endocytic sites is essential for allowing actin polymerization to drive membrane invagination.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
