

Lineage-Specific Profiling Delineates the Emergence and Progression of Naive Pluripotency in Mammalian Embryogenesis
- PMID: 26555056
- PMCID: PMC4643313
- DOI: 10.1016/j.devcel.2015.10.011
Lineage-Specific Profiling Delineates the Emergence and Progression of Naive Pluripotency in Mammalian Embryogenesis
Abstract
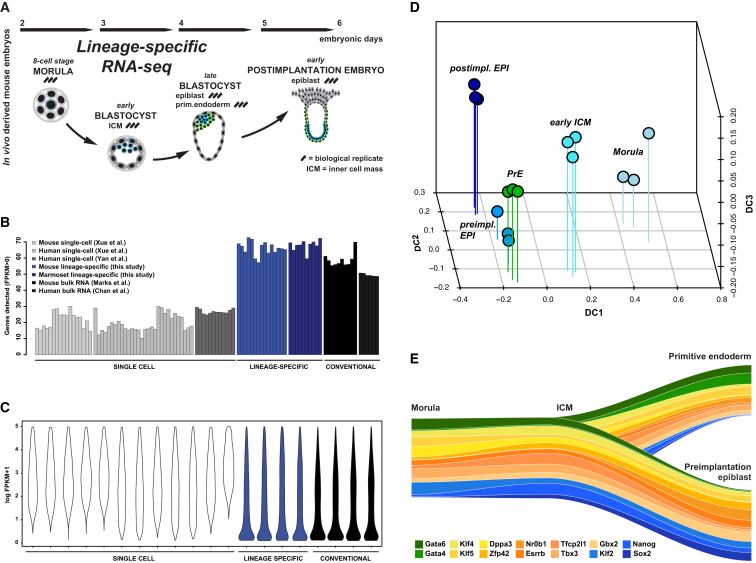
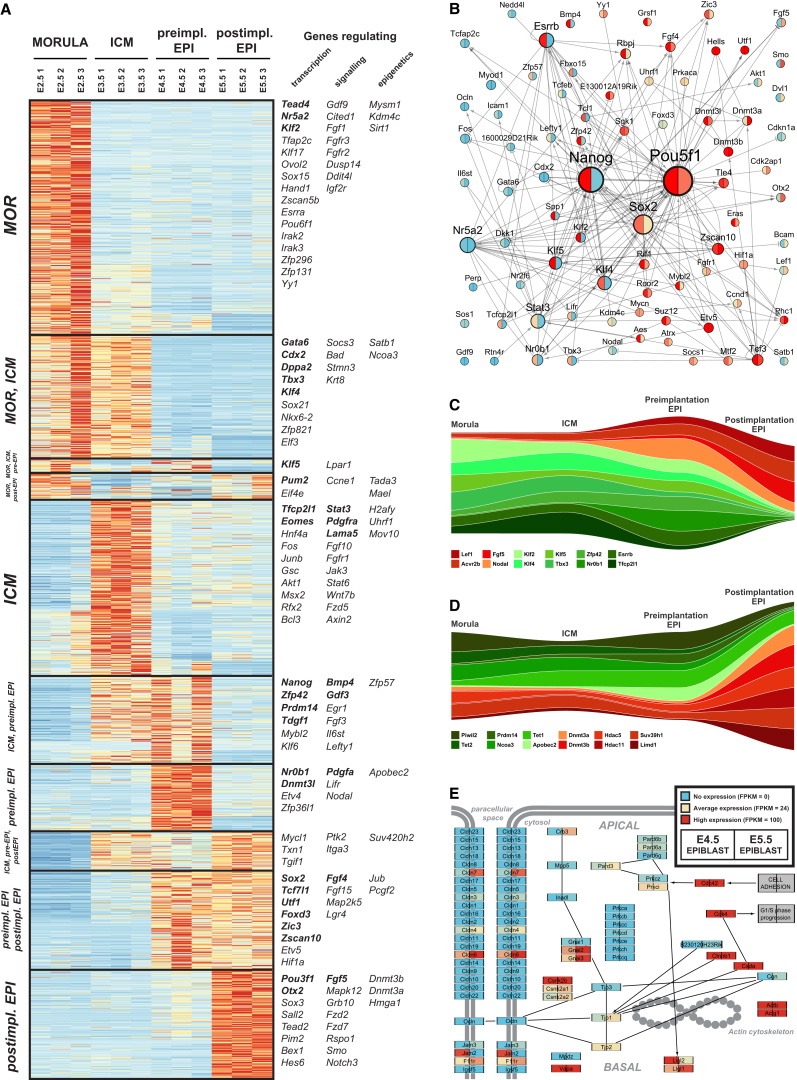
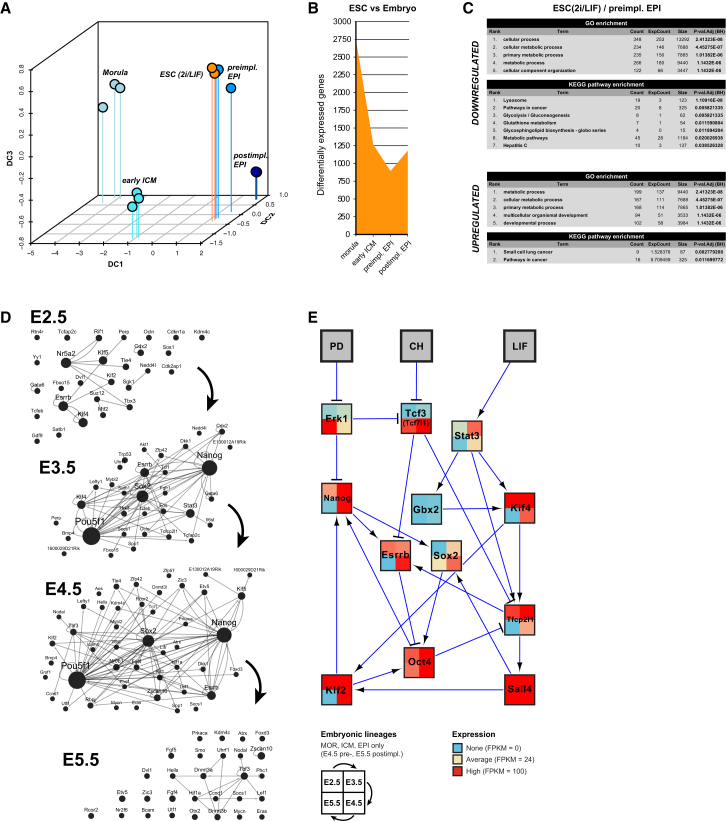
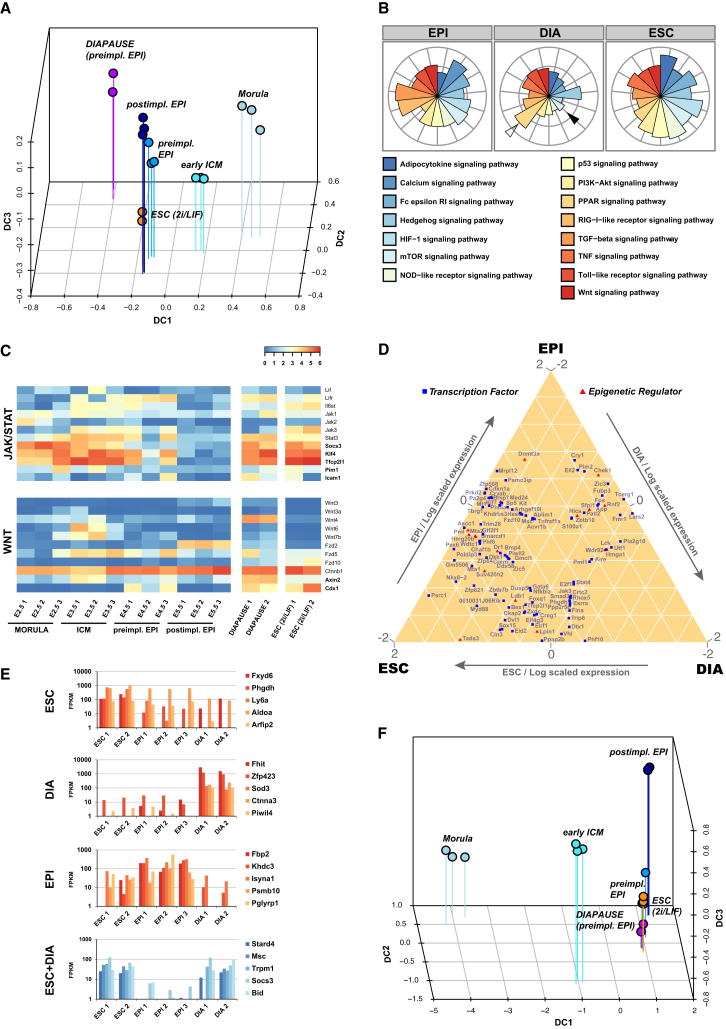
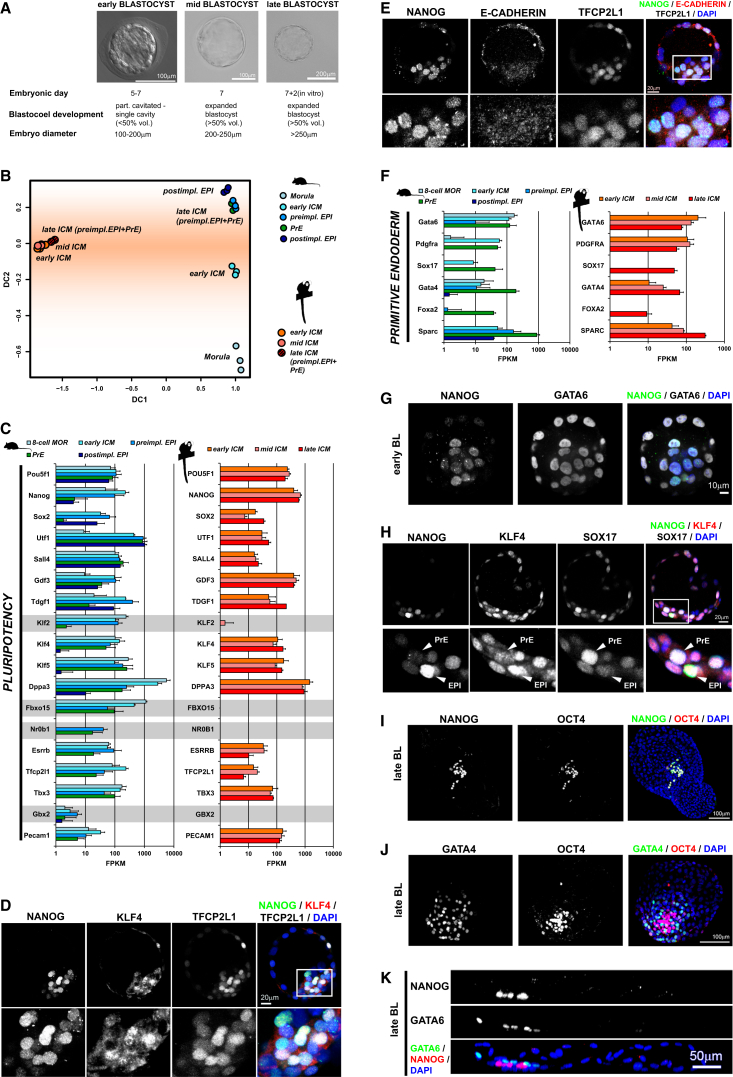
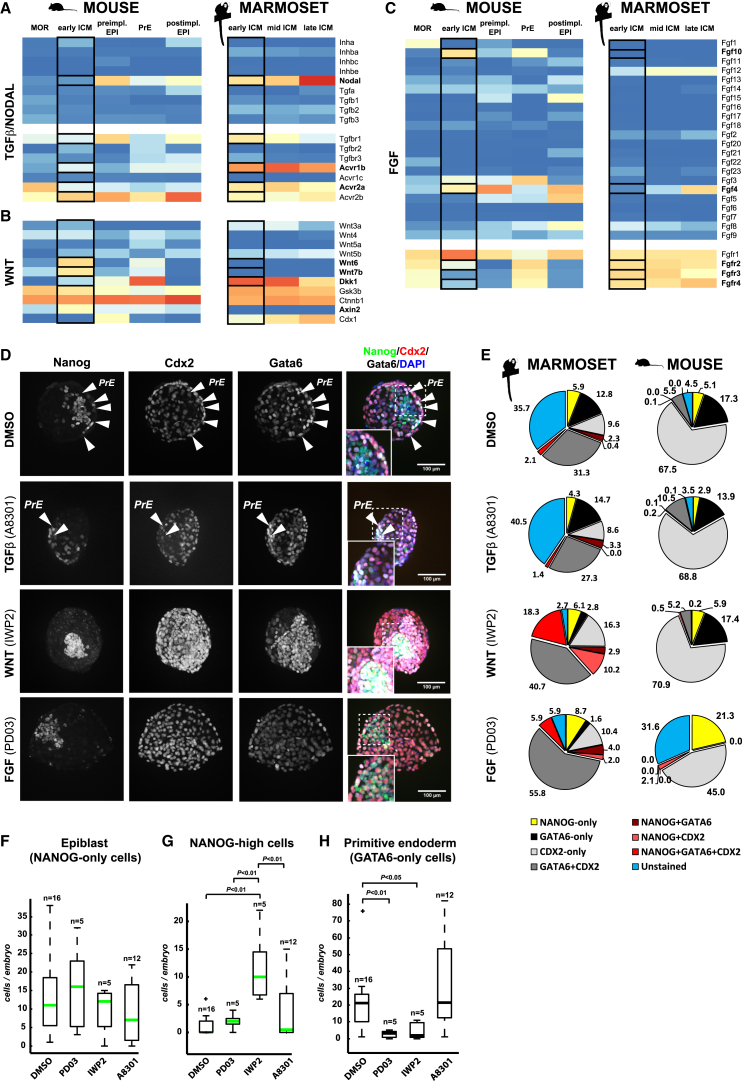
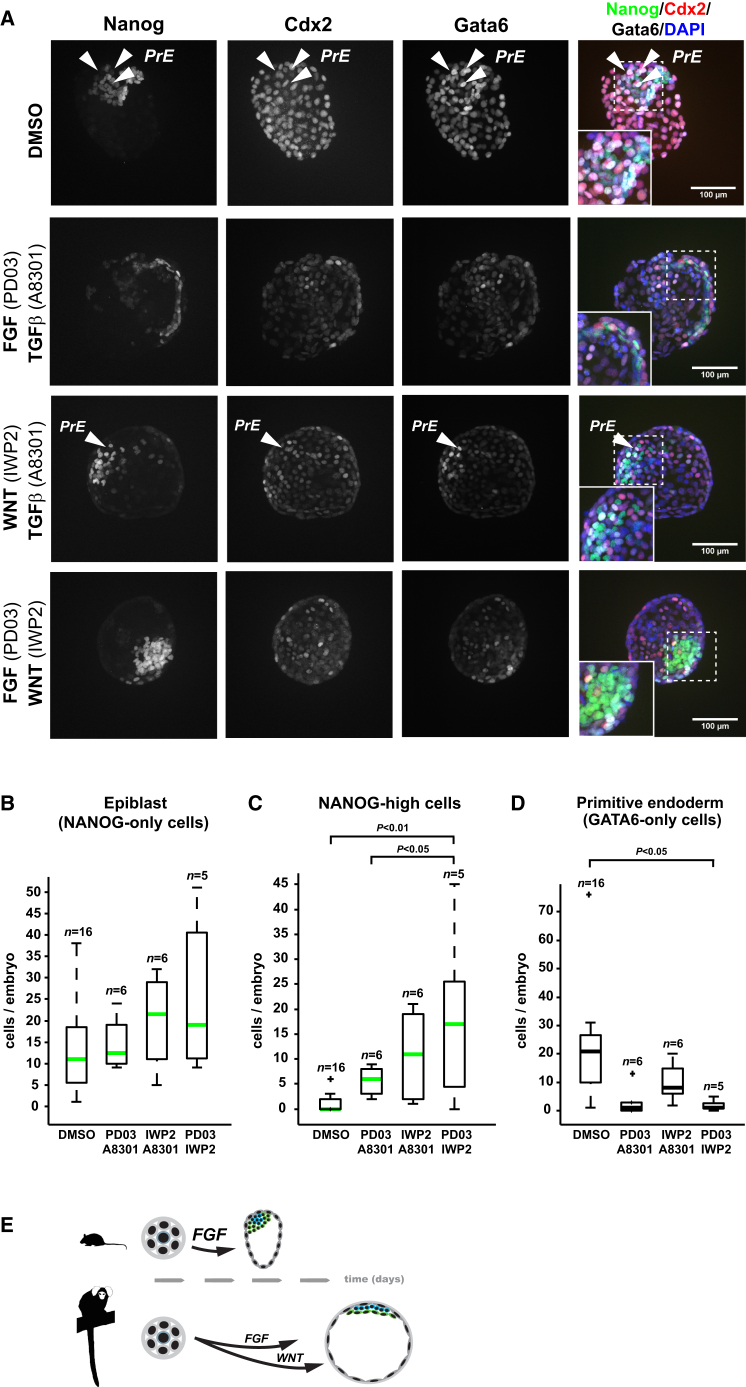
Naive pluripotency is manifest in the preimplantation mammalian embryo. Here we determine transcriptome dynamics of mouse development from the eight-cell stage to postimplantation using lineage-specific RNA sequencing. This method combines high sensitivity and reporter-based fate assignment to acquire the full spectrum of gene expression from discrete embryonic cell types. We define expression modules indicative of developmental state and temporal regulatory patterns marking the establishment and dissolution of naive pluripotency in vivo. Analysis of embryonic stem cells and diapaused embryos reveals near-complete conservation of the core transcriptional circuitry operative in the preimplantation epiblast. Comparison to inner cell masses of marmoset primate blastocysts identifies a similar complement of pluripotency factors but use of alternative signaling pathways. Embryo culture experiments further indicate that marmoset embryos utilize WNT signaling during early lineage segregation, unlike rodents. These findings support a conserved transcription factor foundation for naive pluripotency while revealing species-specific regulatory features of lineage segregation.
Keywords: diapause; embryonic stem cell; inner cell mass; pluripotency; primate.
Copyright © 2015 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
Different Species Choose Their Own Paths to Pluripotency.Dev Cell. 2015 Nov 9;35(3):267-8. doi: 10.1016/j.devcel.2015.10.025. Dev Cell. 2015. PMID: 26555047
References
-
- Adewumi O., Aflatoonian B., Ahrlund-Richter L., Amit M., Andrews P.W., Beighton G., Bello P.A., Benvenisty N., Berry L.S., Bevan S., International Stem Cell Initiative Characterization of human embryonic stem cell lines by the International Stem Cell Initiative. Nat. Biotechnol. 2007;25:803–816. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
