

Meiotic Clade AAA ATPases: Protein Polymer Disassembly Machines
- PMID: 26555750
- PMCID: PMC4860049
- DOI: 10.1016/j.jmb.2015.11.004
Meiotic Clade AAA ATPases: Protein Polymer Disassembly Machines
Abstract
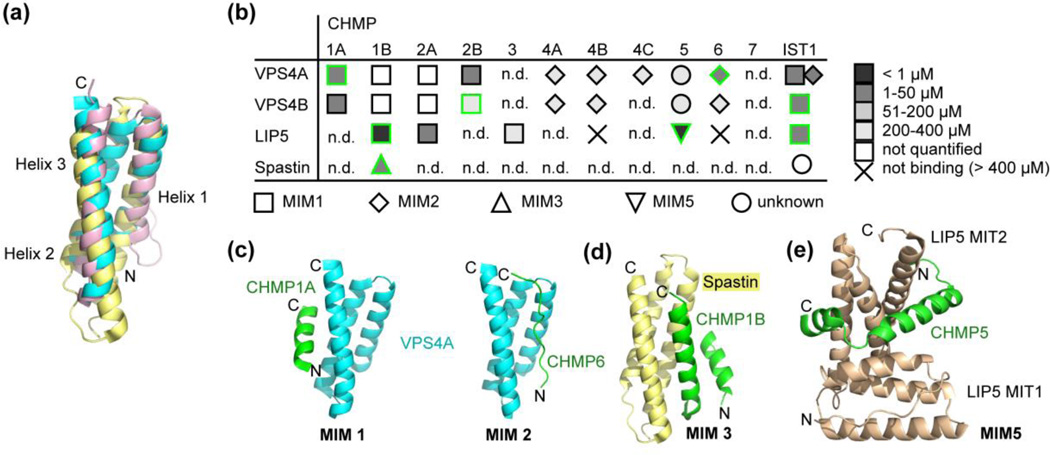
Meiotic clade AAA ATPases (ATPases associated with diverse cellular activities), which were initially grouped on the basis of phylogenetic classification of their AAA ATPase cassette, include four relatively well characterized family members, Vps4, spastin, katanin and fidgetin. These enzymes all function to disassemble specific polymeric protein structures, with Vps4 disassembling the ESCRT-III polymers that are central to the many membrane-remodeling activities of the ESCRT (endosomal sorting complexes required for transport) pathway and spastin, katanin p60 and fidgetin affecting multiple aspects of cellular dynamics by severing microtubules. They share a common domain architecture that features an N-terminal MIT (microtubule interacting and trafficking) domain followed by a single AAA ATPase cassette. Meiotic clade AAA ATPases function as hexamers that can cycle between the active assembly and inactive monomers/dimers in a regulated process, and they appear to disassemble their polymeric substrates by translocating subunits through the central pore of their hexameric ring. Recent studies with Vps4 have shown that nucleotide-induced asymmetry is a requirement for substrate binding to the pore loops and that recruitment to the protein lattice via MIT domains also relieves autoinhibition and primes the AAA ATPase cassettes for substrate binding. The most striking, unifying feature of meiotic clade AAA ATPases may be their MIT domain, which is a module that is found in a wide variety of proteins that localize to ESCRT-III polymers. Spastin also displays an adjacent microtubule binding sequence, and the presence of both ESCRT-III and microtubule binding elements may underlie the recent findings that the ESCRT-III disassembly function of Vps4 and the microtubule-severing function of spastin, as well as potentially katanin and fidgetin, are highly coordinated.
Keywords: ESCRT; Vps4; enzyme mechanism; microtubules; spastin.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures





References
-
- Lupas AN, Martin J. AAA proteins. Curr Opin Struct Biol. 2002;12:746–753. - PubMed
-
- Ammelburg M, Frickey T, Lupas AN. Classification of AAA+ proteins. J Struct Biol. 2006;156:2–11. - PubMed
-
- Frickey T, Lupas AN. Phylogenetic analysis of AAA proteins. J Struct Biol. 2004;146:2–10. - PubMed
-
- Wendler P, Ciniawsky S, Kock M, Kube S. Structure and function of the AAA+ nucleotide binding pocket. Biochim Biophys Acta. 2012;1823:2–14. - PubMed
-
- Neuwald AF, Aravind L, Spouge JL, Koonin EV. AAA+: A class of chaperone-like ATPases associated with the assembly, operation, and disassembly of protein complexes. Genome Res. 1999;9:27–43. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
