

Evolution of the EKA family of powdery mildew avirulence-effector genes from the ORF 1 of a LINE retrotransposon
- PMID: 26556056
- PMCID: PMC4641428
- DOI: 10.1186/s12864-015-2185-x
Evolution of the EKA family of powdery mildew avirulence-effector genes from the ORF 1 of a LINE retrotransposon
Abstract
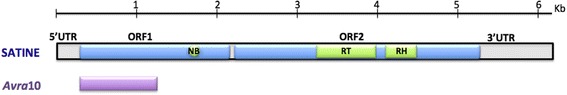
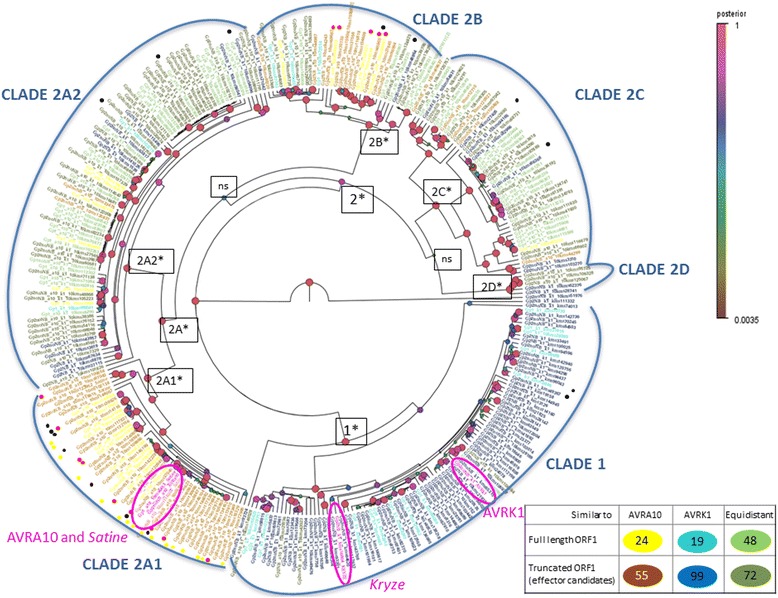
Background: The Avrk1 and Avra10 avirulence (AVR) genes encode effectors that increase the pathogenicity of the fungus Blumeria graminis f.sp. hordei (Bgh), the powdery mildew pathogen, in susceptible barley plants. In resistant barley, MLK1 and MLA10 resistance proteins recognize the presence of AVRK1 and AVRA10, eliciting the hypersensitive response typical of gene for gene interactions. Avrk1 and Avra10 have more than 1350 homologues in Bgh genome, forming the EKA (Effectors homologous to Avr k 1 and Avr a 10) gene family.
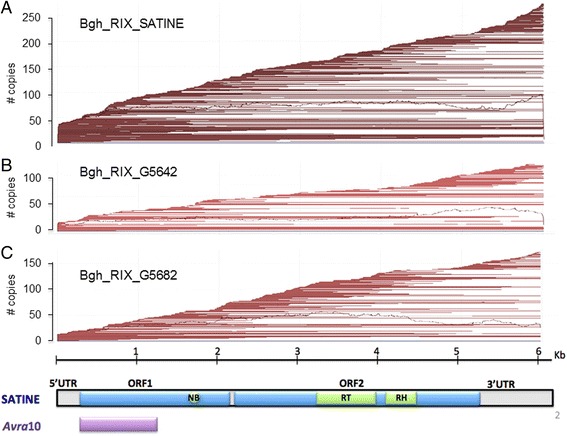
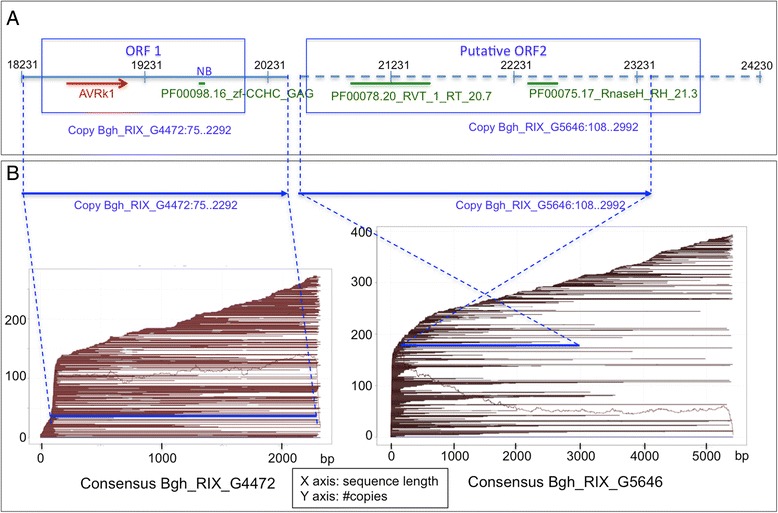
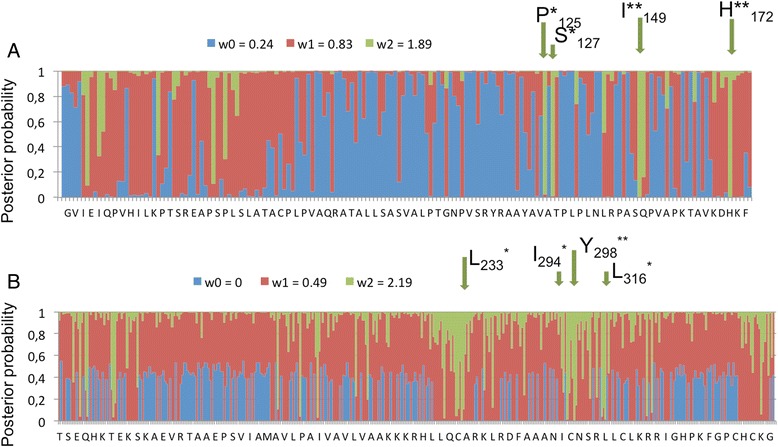
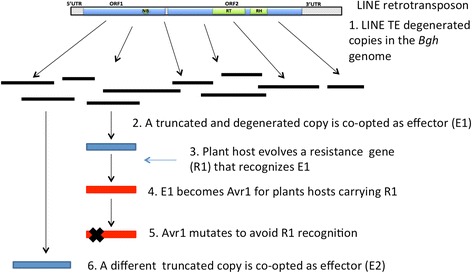
Results: We tested the hypothesis that the EKA family originated from degenerate copies of Class I LINE retrotransposons by analysing the EKA family in the genome of Bgh isolate DH14 with bioinformatic tools specially developed for the analysis of Transposable Elements (TE) in genomes. The Class I LINE retrotransposon copies homologous to Avrk1 and Avra10 represent 6.5 % of the Bgh annotated genome and, among them, we identified 293 AVR/effector candidate genes. We also experimentally identified peptides that indicated the translation of several predicted proteins from EKA family members, which had higher relative abundance in haustoria than in hyphae.
Conclusions: Our analyses indicate that Avrk1 and Avra10 have evolved from part of the ORF1 gene of Class I LINE retrotransposons. The co-option of Avra10 and Avrk1 as effectors from truncated copies of retrotransposons explains the huge number of homologues in Bgh genome that could act as dynamic reservoirs from which new effector genes may evolve. These data provide further evidence for recruitment of retrotransposons in the evolution of new biological functions.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- BB/C506299/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/E000983/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/J/00000605/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/J/0000A252/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
