

Network Events on Multiple Space and Time Scales in Cultured Neural Networks and in a Stochastic Rate Model
- PMID: 26558616
- PMCID: PMC4641680
- DOI: 10.1371/journal.pcbi.1004547
Network Events on Multiple Space and Time Scales in Cultured Neural Networks and in a Stochastic Rate Model
Abstract
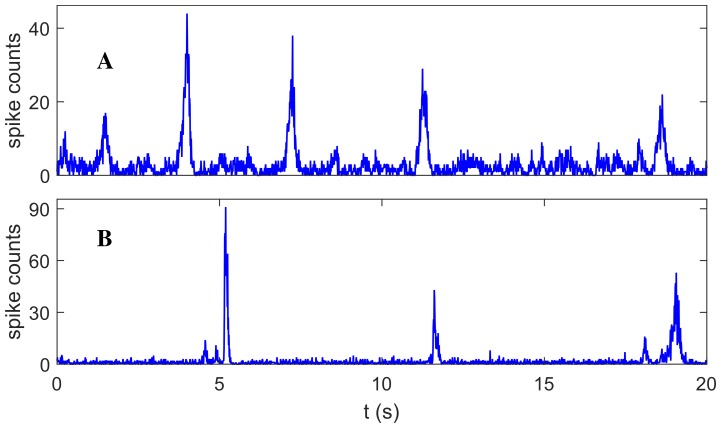
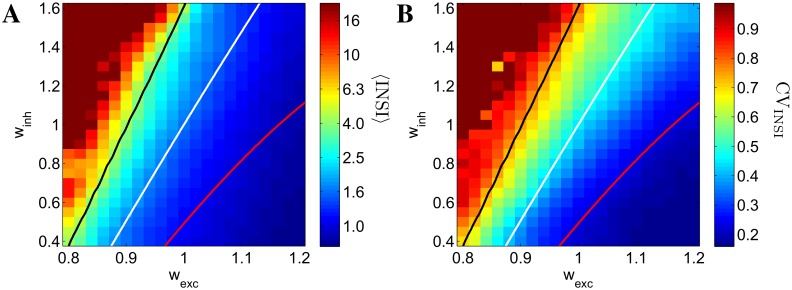
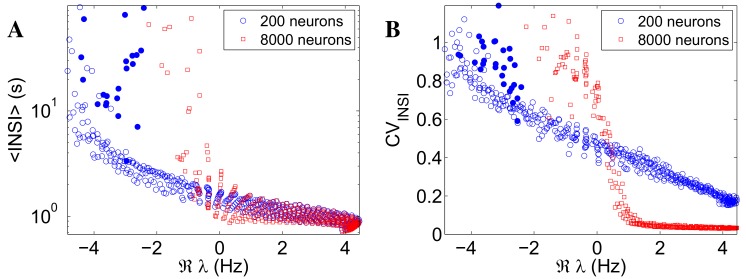
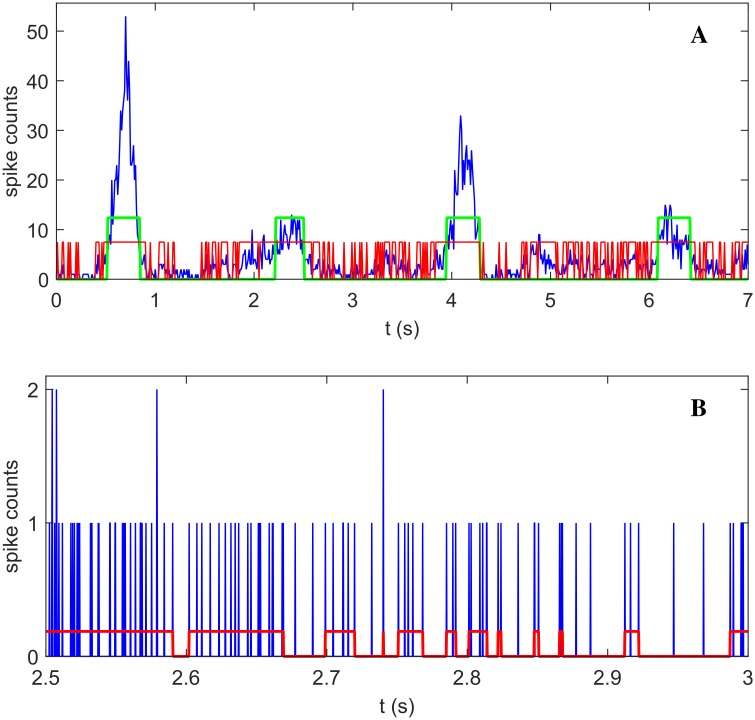
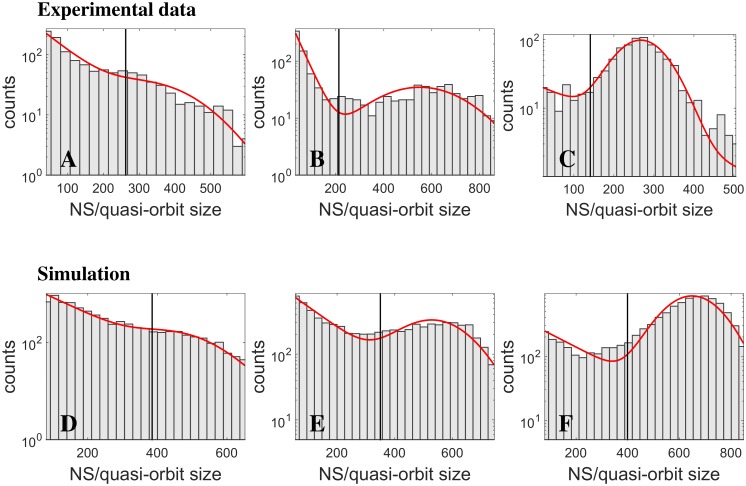
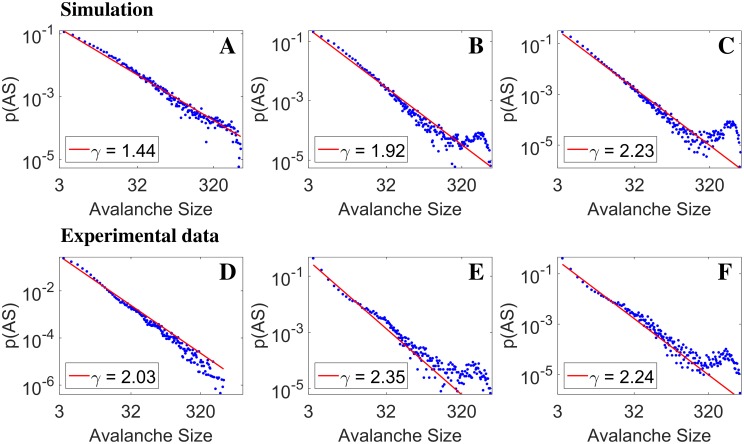
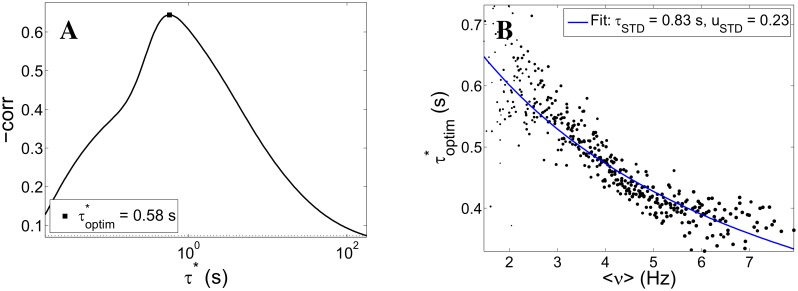
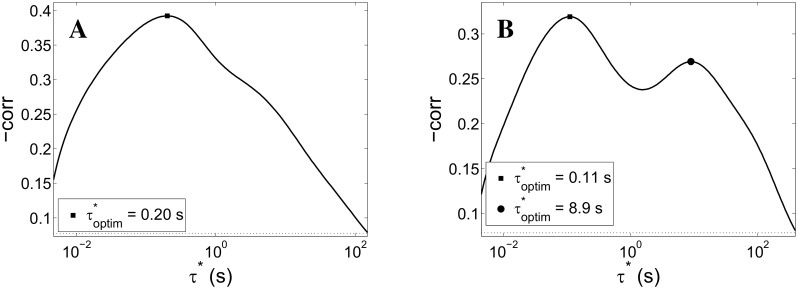
Cortical networks, in-vitro as well as in-vivo, can spontaneously generate a variety of collective dynamical events such as network spikes, UP and DOWN states, global oscillations, and avalanches. Though each of them has been variously recognized in previous works as expression of the excitability of the cortical tissue and the associated nonlinear dynamics, a unified picture of the determinant factors (dynamical and architectural) is desirable and not yet available. Progress has also been partially hindered by the use of a variety of statistical measures to define the network events of interest. We propose here a common probabilistic definition of network events that, applied to the firing activity of cultured neural networks, highlights the co-occurrence of network spikes, power-law distributed avalanches, and exponentially distributed 'quasi-orbits', which offer a third type of collective behavior. A rate model, including synaptic excitation and inhibition with no imposed topology, synaptic short-term depression, and finite-size noise, accounts for all these different, coexisting phenomena. We find that their emergence is largely regulated by the proximity to an oscillatory instability of the dynamics, where the non-linear excitable behavior leads to a self-amplification of activity fluctuations over a wide range of scales in space and time. In this sense, the cultured network dynamics is compatible with an excitation-inhibition balance corresponding to a slightly sub-critical regime. Finally, we propose and test a method to infer the characteristic time of the fatigue process, from the observed time course of the network's firing rate. Unlike the model, possessing a single fatigue mechanism, the cultured network appears to show multiple time scales, signalling the possible coexistence of different fatigue mechanisms.
Conflict of interest statement
Author GG is affiliated with Mperience srl. and this does not alter our adherence to PLOS Computational Biology policies on sharing data and materials. Mperience srl provided support in the form of salaries for author GG, but did not have any additional role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript. The specific roles of these authors are articulated in the ‘author contributions’ section. The other authors have declared that no competing interests exist.
Figures
Similar articles
-
Dynamics of spontaneous activity in random networks with multiple neuron subtypes and synaptic noise : Spontaneous activity in networks with synaptic noise.J Comput Neurosci. 2018 Aug;45(1):1-28. doi: 10.1007/s10827-018-0688-6. Epub 2018 Jun 19. J Comput Neurosci. 2018. PMID: 29923159 Free PMC article.
-
Network bursting dynamics in excitatory cortical neuron cultures results from the combination of different adaptive mechanisms.PLoS One. 2013 Oct 11;8(10):e75824. doi: 10.1371/journal.pone.0075824. eCollection 2013. PLoS One. 2013. PMID: 24146781 Free PMC article.
-
Self-organization and neuronal avalanches in networks of dissociated cortical neurons.Neuroscience. 2008 Jun 2;153(4):1354-69. doi: 10.1016/j.neuroscience.2008.03.050. Epub 2008 Mar 29. Neuroscience. 2008. PMID: 18448256
-
Neural dynamics in cortical networks--precision of joint-spiking events.Novartis Found Symp. 2001;239:193-204; discussion 204-7, 234-40. doi: 10.1002/0470846674.ch15. Novartis Found Symp. 2001. PMID: 11529312 Review.
-
Space-Time Dynamics of Membrane Currents Evolve to Shape Excitation, Spiking, and Inhibition in the Cortex at Small and Large Scales.Neuron. 2017 Jun 7;94(5):934-942. doi: 10.1016/j.neuron.2017.04.038. Neuron. 2017. PMID: 28595049 Review.
Cited by
-
Interleaving asynchronous and synchronous activity in balanced cortical networks with short-term synaptic depression.bioRxiv [Preprint]. 2025 Mar 18:2025.03.13.643074. doi: 10.1101/2025.03.13.643074. bioRxiv. 2025. PMID: 40166256 Free PMC article. Preprint.
-
Low-dimensional spike rate models derived from networks of adaptive integrate-and-fire neurons: Comparison and implementation.PLoS Comput Biol. 2017 Jun 23;13(6):e1005545. doi: 10.1371/journal.pcbi.1005545. eCollection 2017 Jun. PLoS Comput Biol. 2017. PMID: 28644841 Free PMC article.
-
Unique Features of Network Bursts Emerge From the Complex Interplay of Excitatory and Inhibitory Receptors in Rat Neocortical Networks.Front Cell Neurosci. 2019 Sep 6;13:377. doi: 10.3389/fncel.2019.00377. eCollection 2019. Front Cell Neurosci. 2019. PMID: 31555093 Free PMC article.
-
Unstructured network topology begets order-based representation by privileged neurons.Biol Cybern. 2020 Feb;114(1):113-135. doi: 10.1007/s00422-020-00819-9. Epub 2020 Feb 27. Biol Cybern. 2020. PMID: 32107622 Free PMC article.
-
Mesoscopic description of hippocampal replay and metastability in spiking neural networks with short-term plasticity.PLoS Comput Biol. 2022 Dec 22;18(12):e1010809. doi: 10.1371/journal.pcbi.1010809. eCollection 2022 Dec. PLoS Comput Biol. 2022. PMID: 36548392 Free PMC article.
References
-
- Park IPI, Xu DXD, DeMarse TB, Principe JC (2006). Modeling of Synchronized Burst in Dissociated Cortical Tissue: An Exploration of Parameter Space. 10.1109/IJCNN.2006.246734 - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
