

Seven New Complete Plastome Sequences Reveal Rampant Independent Loss of the ndh Gene Family across Orchids and Associated Instability of the Inverted Repeat/Small Single-Copy Region Boundaries
- PMID: 26558895
- PMCID: PMC4641739
- DOI: 10.1371/journal.pone.0142215
Seven New Complete Plastome Sequences Reveal Rampant Independent Loss of the ndh Gene Family across Orchids and Associated Instability of the Inverted Repeat/Small Single-Copy Region Boundaries
Abstract
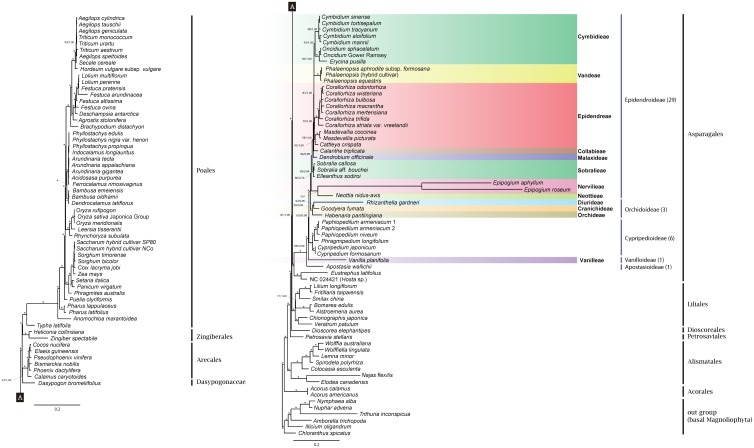
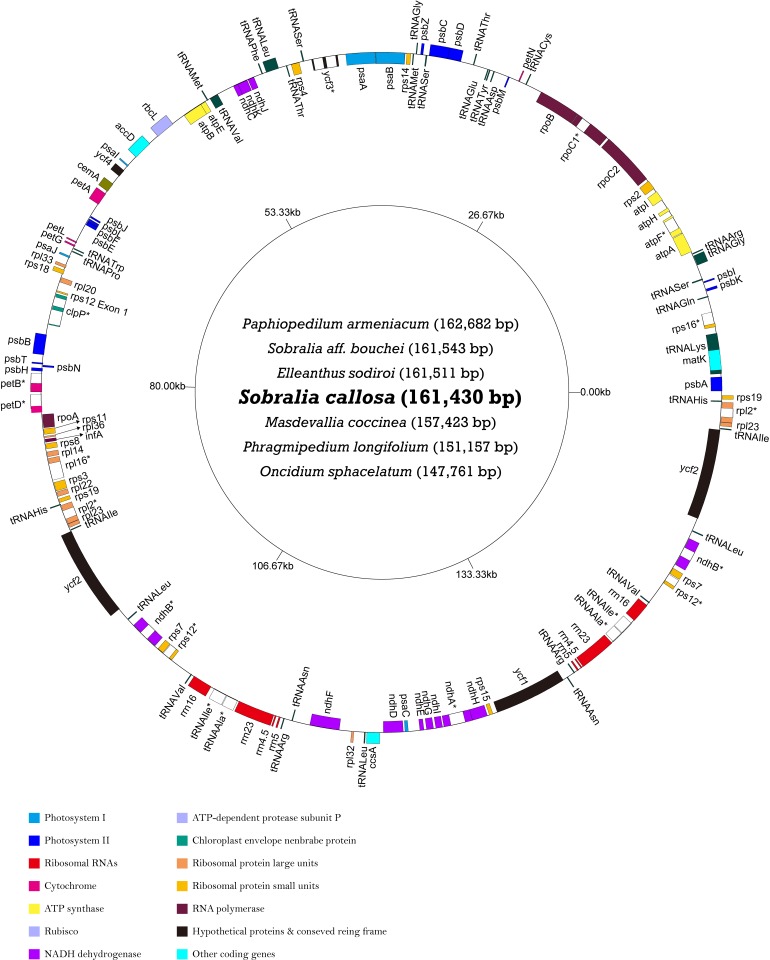
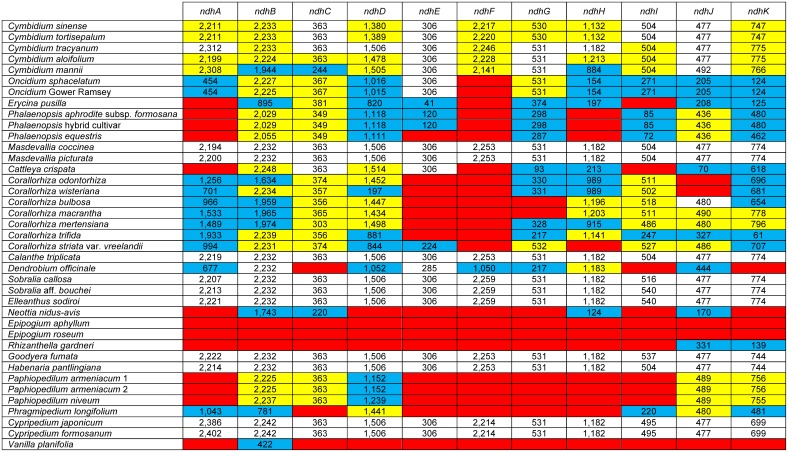
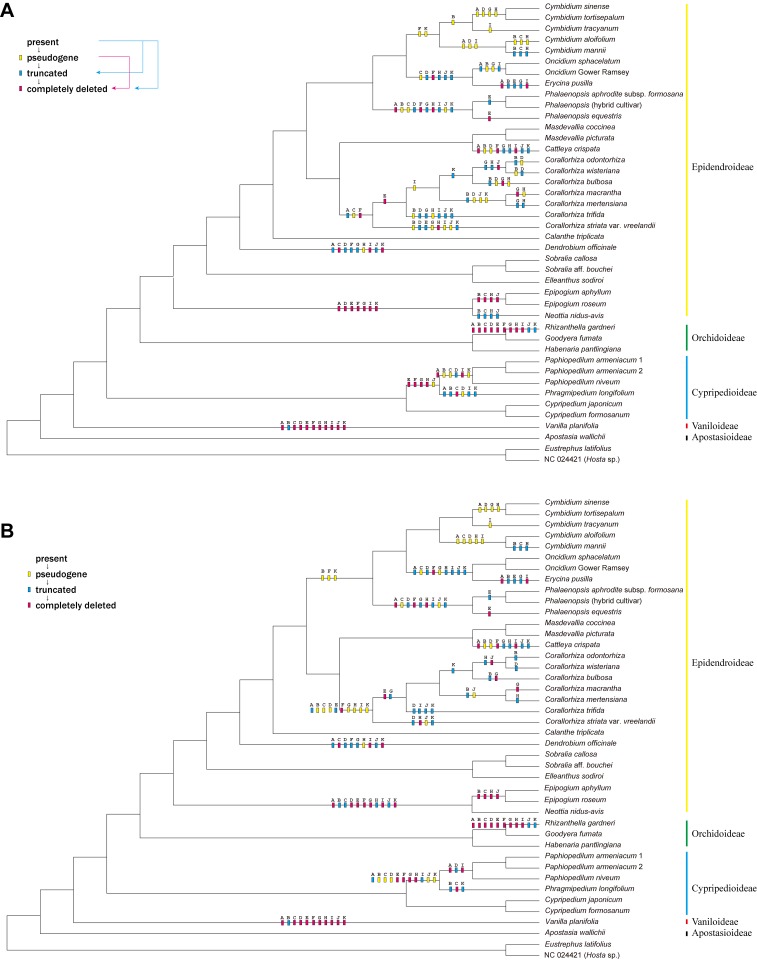
Earlier research has revealed that the ndh loci have been pseudogenized, truncated, or deleted from most orchid plastomes sequenced to date, including in all available plastomes of the two most species-rich subfamilies, Orchidoideae and Epidendroideae. This study sought to resolve deeper-level phylogenetic relationships among major orchid groups and to refine the history of gene loss in the ndh loci across orchids. The complete plastomes of seven orchids, Oncidium sphacelatum (Epidendroideae), Masdevallia coccinea (Epidendroideae), Sobralia callosa (Epidendroideae), Sobralia aff. bouchei (Epidendroideae), Elleanthus sodiroi (Epidendroideae), Paphiopedilum armeniacum (Cypripedioideae), and Phragmipedium longifolium (Cypripedioideae) were sequenced and analyzed in conjunction with all other available orchid and monocot plastomes. Most ndh loci were found to be pseudogenized or lost in Oncidium, Paphiopedilum and Phragmipedium, but surprisingly, all ndh loci were found to retain full, intact reading frames in Sobralia, Elleanthus and Masdevallia. Character mapping suggests that the ndh genes were present in the common ancestor of orchids but have experienced independent, significant losses at least eight times across four subfamilies. In addition, ndhF gene loss was correlated with shifts in the position of the junction of the inverted repeat (IR) and small single-copy (SSC) regions. The Orchidaceae have unprecedented levels of homoplasy in ndh gene presence/absence, which may be correlated in part with the unusual life history of orchids. These results also suggest that ndhF plays a role in IR/SSC junction stability.
Conflict of interest statement
Figures

References
-
- Govaerts R, Dransfield J, Zona S, Hodel D, Henderson A. World Checklist of Orchidaceae. Facilitated by the Royal Botanic Gardens, Kew. Available: http://apps.kew.org/wcsp/.
-
- Cameron KM, Chase MW, Whitten WM, Kores PJ, Jarrell DC, Albert VA, et al. A phylogenetic analysis of the Orchidaceae: evidence from rbcL nucleotide sequences. Am J Bot. 1999;86(2): 208–224. - PubMed
-
- Albert V. Cladistic relationships of the slipper orchids (Cypripedioideae: Orchidaceae) from congruent morphological and molecular data. Lindleyana. 1994;9(2): 115–132.
-
- Yukawa T, Ohba H, Cameron KM, Chase MW. Chloroplast DNA phylogeny of subtribe Dendrobiinae (Orchidaceae): Insights from a combined analysis based on rbcL sequences and restriction site variation. Journal of Plant Research. 1996;109(1094): 169–176. 10.1007/Bf02344542 - DOI
-
- Kores P, Cameron K, Molvray M, Chase M. The phylogenetic relationships of Orchidoideae and Spiranthoideae (Orchidaceae) as inferred from rbcL plastid sequences. Lindleyana. 1997;12(1): 1–11.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
