

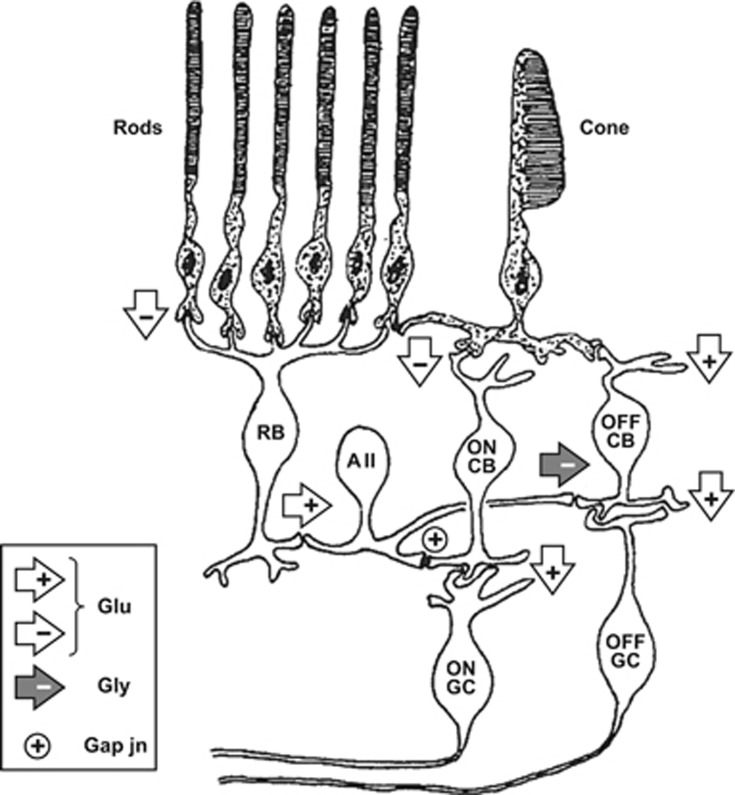
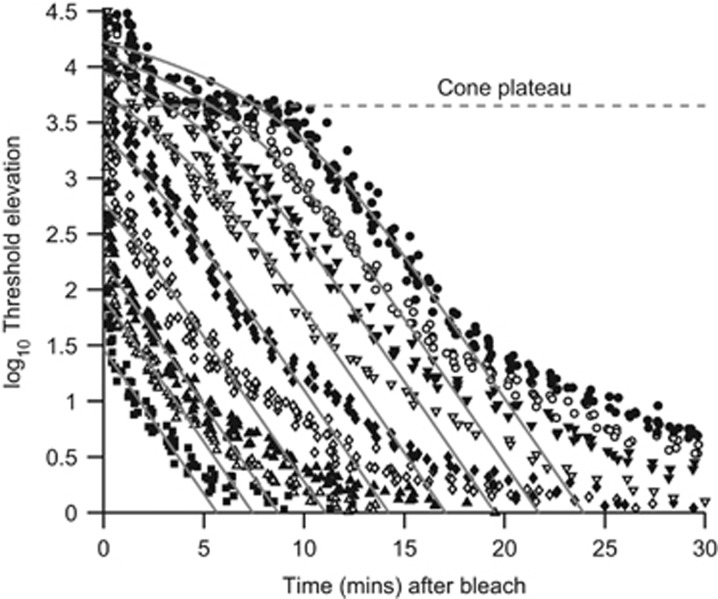
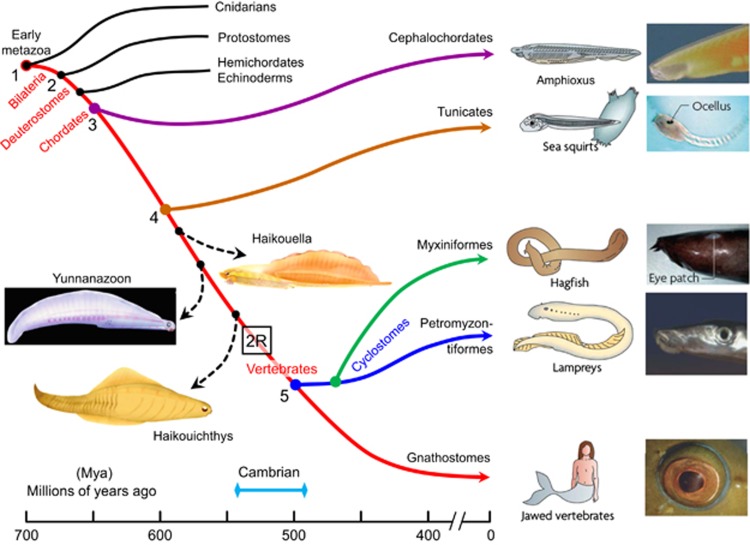
Why rods and cones?
- PMID: 26563661
- PMCID: PMC4763127
- DOI: 10.1038/eye.2015.236
Why rods and cones?
Abstract
Under twenty-first-century metropolitan conditions, almost all of our vision is mediated by cones and the photopic system, yet cones make up barely 5% of our retinal photoreceptors. This paper looks at reasons why we additionally possess rods and a scotopic system, and asks why rods comprise 95% of our retinal photoreceptors. It considers the ability of rods to reliably signal the arrival of individual photons of light, as well as the ability of the retina to process these single-photon signals, and it discusses the advantages that accrue. Drawbacks in the arrangement, including the very slow dark adaptation of scotopic vision, are also considered. Finally, the timing of the evolution of cone and rod photoreceptors, the retina, and the camera-style eye is summarised.
Figures
References
-
- 1Østerberg G. Topography of the layer of rods and cones in the human retina. Acta Ophthalmol 1935; 13(S6): 1–103.
-
- 2Curcio CA, Sloan KR, Kalina RE, Hendrickson AE. Human photoreceptor topography. J Comp Neurol 1990; 292: 497–523. - PubMed
-
- 3Curcio CA, Hendrickson AE. Organization and development of the primate photoreceptor mosaic. Prog Retinal Res 1991; 10: 89–120.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
