

Reactivation of autophagy by spermidine ameliorates the myopathic defects of collagen VI-null mice
- PMID: 26565691
- PMCID: PMC4835186
- DOI: 10.1080/15548627.2015.1108508
Reactivation of autophagy by spermidine ameliorates the myopathic defects of collagen VI-null mice
Abstract
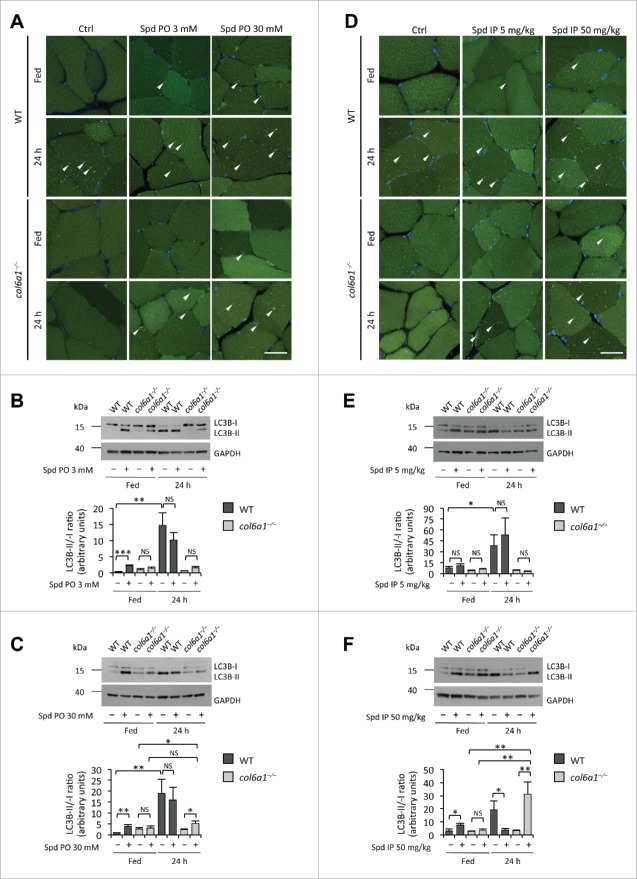
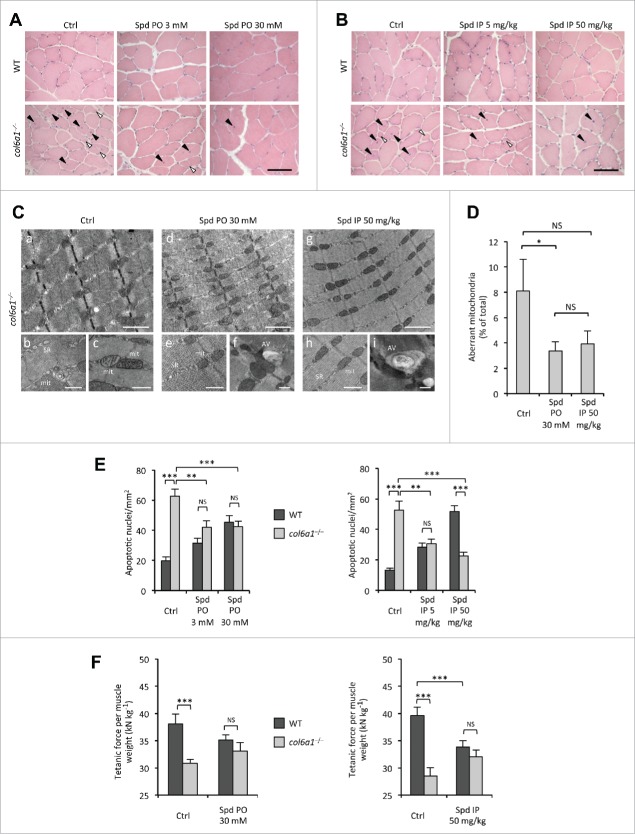
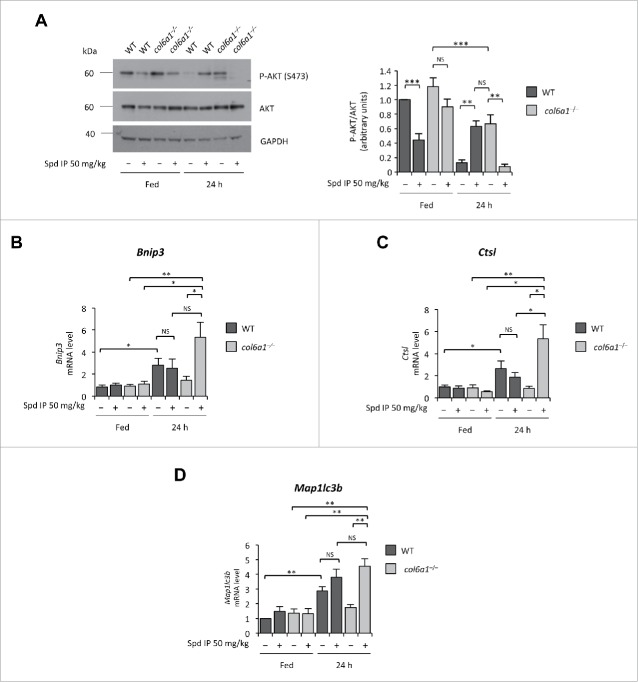
Autophagy is a self-degradative process responsible for the clearance of damaged or unnecessary cellular components. We have previously found that persistence of dysfunctional organelles due to autophagy failure is a key event in the pathogenesis of COL6/collagen VI-related myopathies, and have demonstrated that reactivation of a proper autophagic flux rescues the muscle defects of Col6a1-null (col6a1(-/-)) mice. Here we show that treatment with spermidine, a naturally occurring nontoxic autophagy inducer, is beneficial for col6a1(-/-) mice. Systemic administration of spermidine in col6a1(-/-) mice reactivated autophagy in a dose-dependent manner, leading to a concurrent amelioration of the histological and ultrastructural muscle defects. The beneficial effects of spermidine, together with its being easy to administer and the lack of overt side effects, open the field for the design of novel nutraceutical strategies for the treatment of muscle diseases characterized by autophagy impairment.
Keywords: autophagy; collagen VI; muscular dystrophies; skeletal muscle; spermidine.
Figures
References
-
- Mizushima N. The pleiotropic role of autophagy: from protein metabolism to bactericide. Cell Death Differ 2005; 12 Suppl 2:1535-41; PMID:16247501; http://dx.doi.org/ 10.1038/sj.cdd.4401728 - DOI - PubMed
-
- Cecconi F, Levine B. The role of autophagy in mammalian development: cell makeover rather than cell death. Dev Cell 2008; 15:344-57; PMID:18804433; http://dx.doi.org/ 10.1016/j.devcel.2008.08.012 - DOI - PMC - PubMed
-
- Levine B, Kroemer G. Autophagy in the pathogenesis of disease. Cell 2008; 132:27-42; PMID:18191218; http://dx.doi.org/ 10.1016/j.cell.2007.12.018 - DOI - PMC - PubMed
-
- Rogov V, Dötsch V, Johansen T, Kirkin V. Interactions between autophagy receptors and ubiquitin-like proteins form the molecular basis for selective autophagy. Mol Cell 2014; 53:167-78; PMID:24462201; http://dx.doi.org/ 10.1016/j.molcel.2013.12.014 - DOI - PubMed
-
- Sandri M, Coletto L, Grumati P, Bonaldo P. Misregulation of autophagy and protein degradation systems in myopathies and muscular dystrophies. J Cell Sci 2013; 126:5325-33; PMID:24293330; http://dx.doi.org/ 10.1242/jcs.114041 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials