

Proteasome Inhibition Suppresses Dengue Virus Egress in Antibody Dependent Infection
- PMID: 26565697
- PMCID: PMC4643959
- DOI: 10.1371/journal.pntd.0004058
Proteasome Inhibition Suppresses Dengue Virus Egress in Antibody Dependent Infection
Abstract
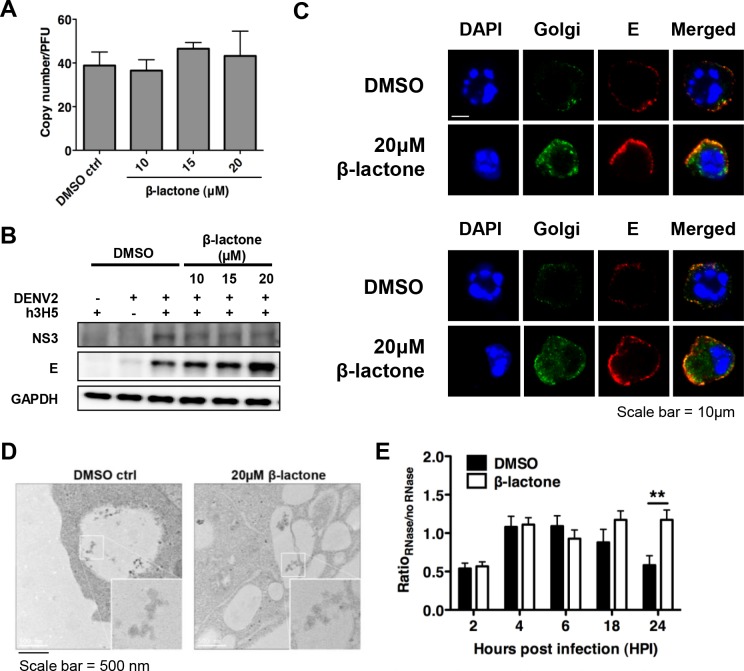
The mosquito-borne dengue virus (DENV) is a cause of significant global health burden, with an estimated 390 million infections occurring annually. However, no licensed vaccine or specific antiviral treatment for dengue is available. DENV interacts with host cell factors to complete its life cycle although this virus-host interplay remains to be fully elucidated. Many studies have identified the ubiquitin proteasome pathway (UPP) to be important for successful DENV production, but how the UPP contributes to DENV life cycle as host factors remains ill defined. We show here that proteasome inhibition decouples infectious virus production from viral RNA replication in antibody-dependent infection of THP-1 cells. Molecular and imaging analyses in β-lactone treated THP-1 cells suggest that proteasome function does not prevent virus assembly but rather DENV egress. Intriguingly, the licensed proteasome inhibitor, bortezomib, is able to inhibit DENV titers at low nanomolar drug concentrations for different strains of all four serotypes of DENV in primary monocytes. Furthermore, bortezomib treatment of DENV-infected mice inhibited the spread of DENV in the spleen as well as the overall pathological changes. Our findings suggest that preventing DENV egress through proteasome inhibition could be a suitable therapeutic strategy against dengue.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
-
- Capeding MR, Tran NH, Hadinegoro SR, Ismail HI, Chotpitayasunondh T, et al. (2014) Clinical efficacy and safety of a novel tetravalent dengue vaccine in healthy children in Asia: a phase 3, randomised, observer-masked, placebo-controlled trial. Lancet 384: 1358–1365. 10.1016/S0140-6736(14)61060-6 - DOI - PubMed
-
- Guzman MG, Kouri G, Valdes L, Bravo J, Vazquez S, et al. (2002) Enhanced severity of secondary dengue-2 infections: death rates in 1981 and 1997 Cuban outbreaks. Rev Panam Salud Publica 11: 223–227. - PubMed
-
- Kliks SC, Nimmanitya S, Nisalak A, Burke DS (1988) Evidence that maternal dengue antibodies are important in the development of dengue hemorrhagic fever in infants. Am J Trop Med Hyg 38: 411–419. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
