

Autophagy contributes to regulation of the hypoxia response during submergence in Arabidopsis thaliana
- PMID: 26566261
- PMCID: PMC4835207
- DOI: 10.1080/15548627.2015.1112483
Autophagy contributes to regulation of the hypoxia response during submergence in Arabidopsis thaliana
Abstract
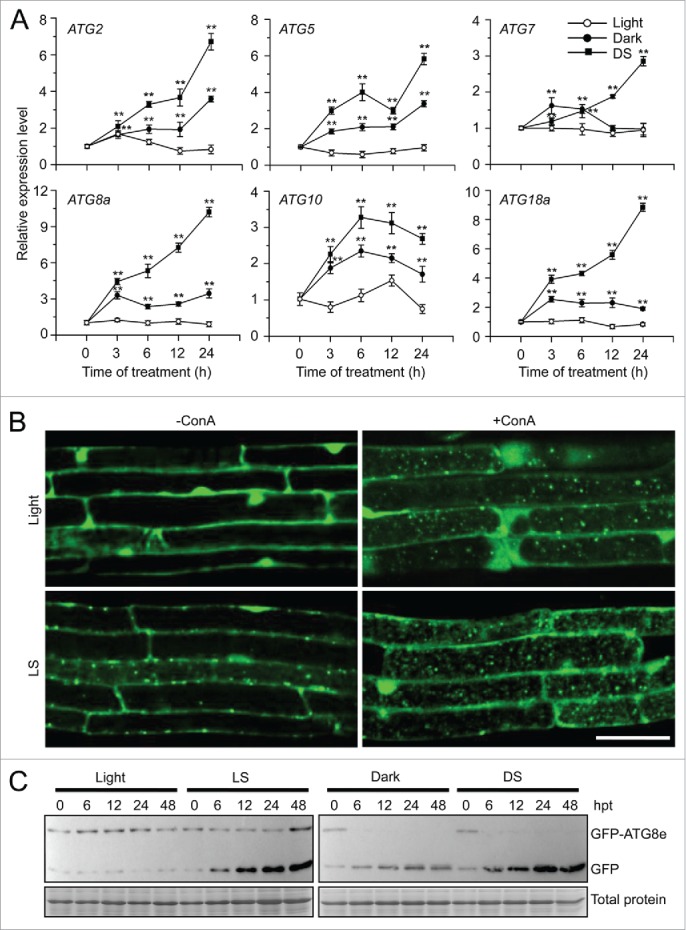
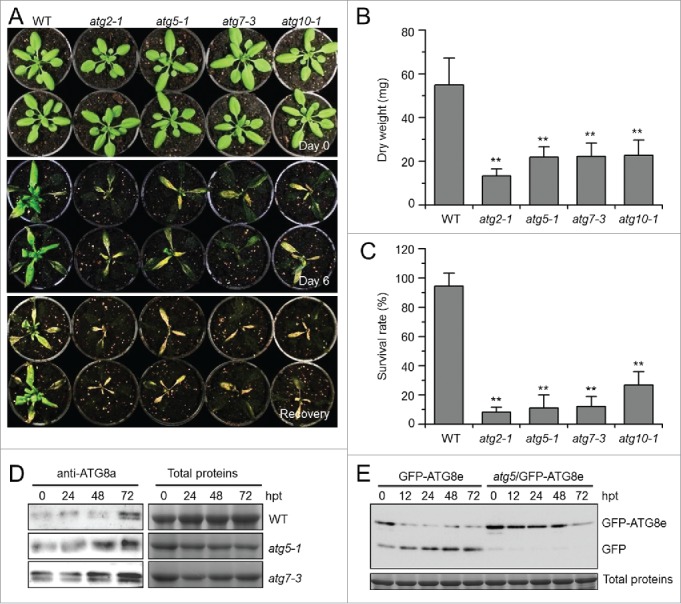
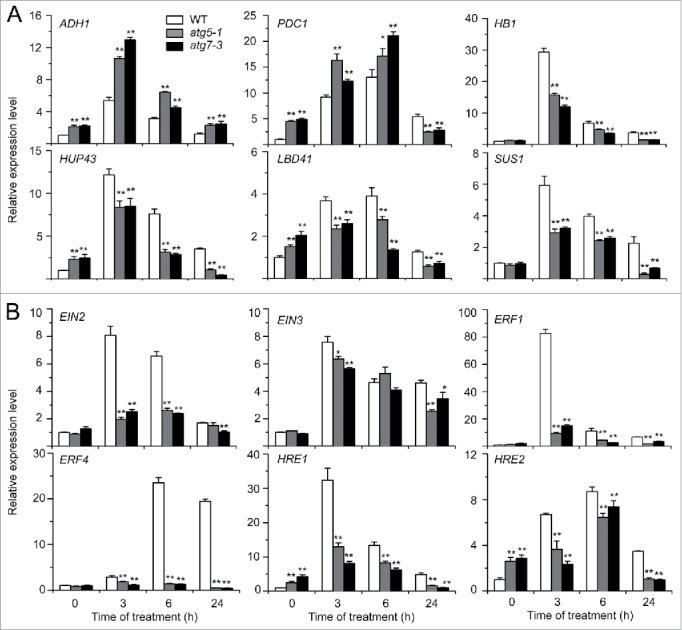
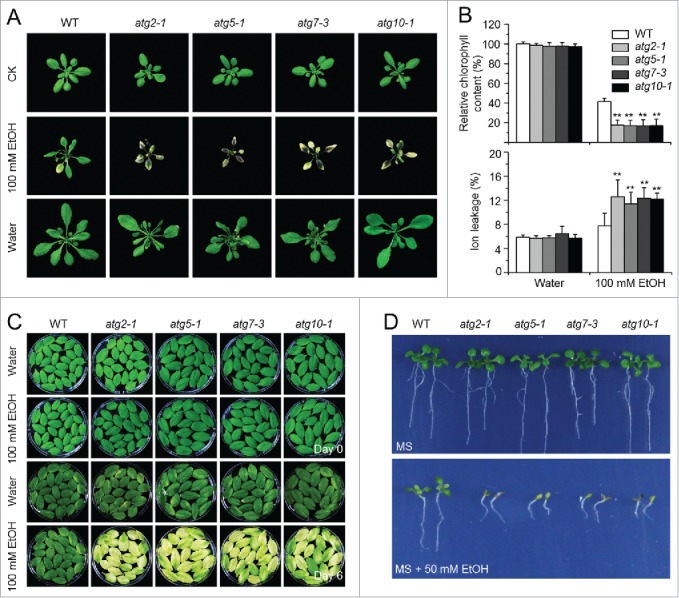
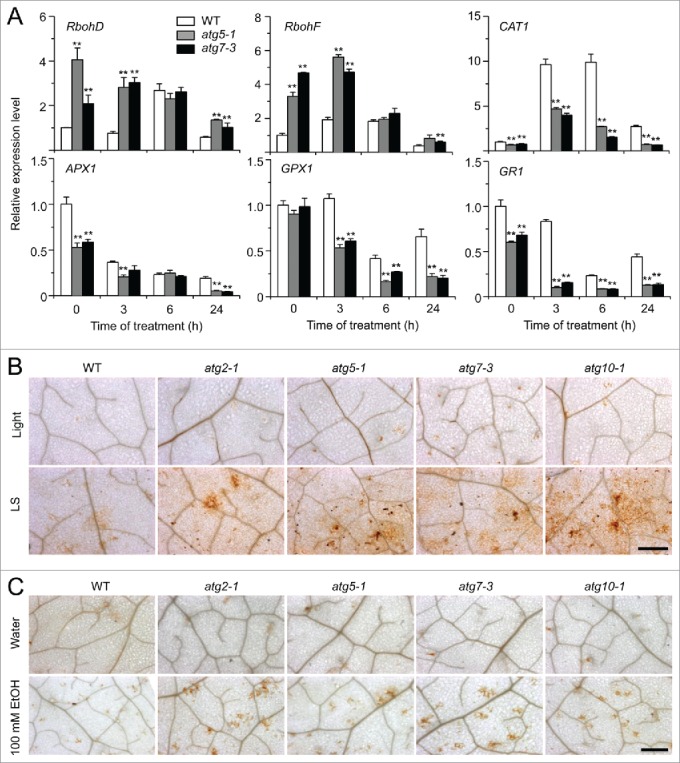
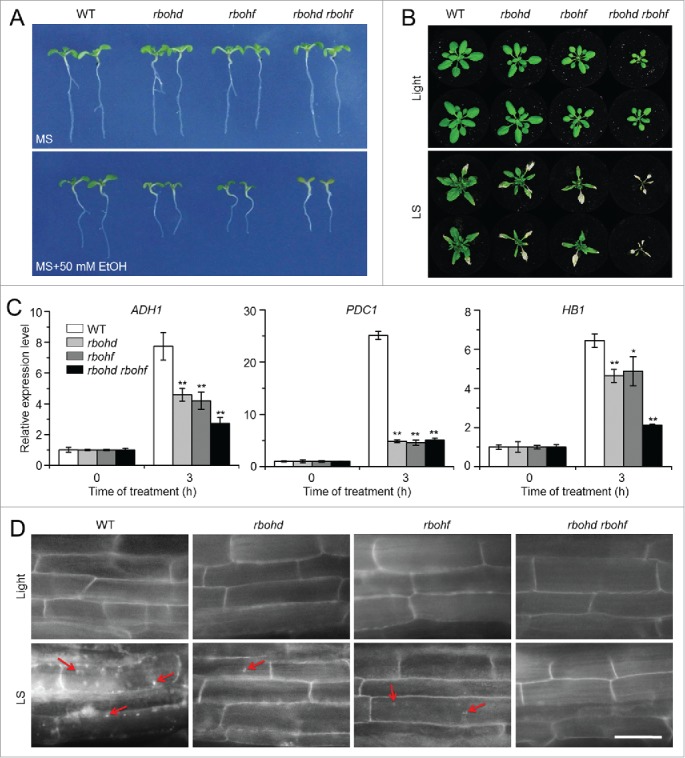
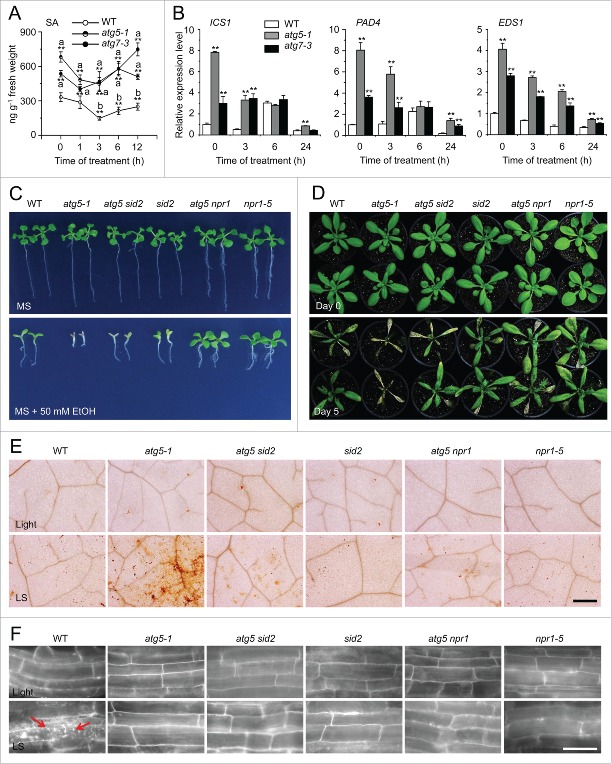
Autophagy involves massive degradation of intracellular components and functions as a conserved system that helps cells to adapt to adverse conditions. In mammals, hypoxia rapidly stimulates autophagy as a cell survival response. Here, we examine the function of autophagy in the regulation of the plant response to submergence, an abiotic stress that leads to hypoxia and anaerobic respiration in plant cells. In Arabidopsis thaliana, submergence induces the transcription of autophagy-related (ATG) genes and the formation of autophagosomes. Consistent with this, the autophagy-defective (atg) mutants are hypersensitive to submergence stress and treatment with ethanol, the end product of anaerobic respiration. Upon submergence, the atg mutants have increased levels of transcripts of anaerobic respiration genes (alcohol dehydrogenase 1, ADH1 and pyruvate decarboxylase 1, PDC1), but reduced levels of transcripts of other hypoxia- and ethylene-responsive genes. Both submergence and ethanol treatments induce the accumulation of reactive oxygen species (ROS) in the rosettes of atg mutants more than in the wild type. Moreover, the production of ROS by the nicotinamide adenine dinucleotide phosphate (NADPH) oxidases is necessary for plant tolerance to submergence and ethanol, submergence-induced expression of ADH1 and PDC1, and activation of autophagy. The submergence- and ethanol-sensitive phenotypes in the atg mutants depend on a complete salicylic acid (SA) signaling pathway. Together, our findings demonstrate that submergence-induced autophagy functions in the hypoxia response in Arabidopsis by modulating SA-mediated cellular homeostasis.
Keywords: autophagy; ethanol; hypoxia; reactive oxygen species; submergence.
Figures
References
-
- Bailey-Serres J, Fukao T, Gibbs DJ, Holdsworth MJ, Lee SC, Licausi F, Perata P, Voesenek LACJ, van Dongen JT. Making sense of low oxygen sensing. Trends Plant Sci 2012; 17:129-38; PMID:22280796; http://dx.doi.org/ 10.1016/j.tplants.2011.12.004 - DOI - PubMed
-
- Bailey-Serres J, Voesenek LACJ. Flooding stress: acclimations and genetic diversity. Annu Rev Plant Biol 2008; 59:313-39; PMID:18444902; http://dx.doi.org/ 10.1146/annurev.arplant.59.032607.092752 - DOI - PubMed
-
- Voesenek LACJ, Sasidharan R. Ethylene– and oxygen signaling– drive plant survival during flooding. Plant Biol 2013; 15:426-35; PMID:23574304; http://dx.doi.org/ 10.1111/plb.12014 - DOI - PubMed
-
- Geigenberger P. Response of plant metabolism to too little oxygen. Curr. Opin. Plant Biol 2003; 6:247-56; PMID:12753974; http://dx.doi.org/ 10.1016/S1369-5266(03)00038-4 - DOI - PubMed
-
- Perata P, Voesenek LACJ. Submergence tolerance in rice requires Sub1A, an ethylene-response-factor-like gene. Trends Plant Sci 2007; 12:43-6; PMID:17208508; http://dx.doi.org/ 10.1016/j.tplants.2006.12.005 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases