

Evolution of translation machinery in recoded bacteria enables multi-site incorporation of nonstandard amino acids
- PMID: 26571098
- PMCID: PMC4784704
- DOI: 10.1038/nbt.3372
Evolution of translation machinery in recoded bacteria enables multi-site incorporation of nonstandard amino acids
Abstract
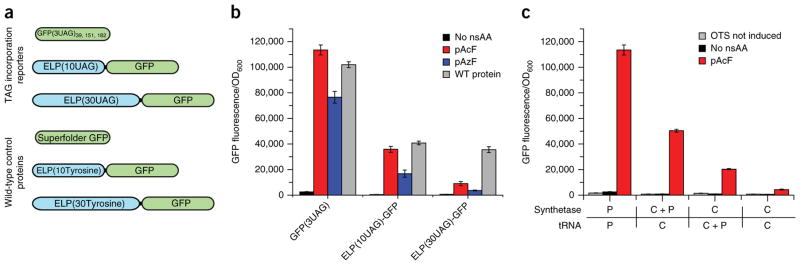
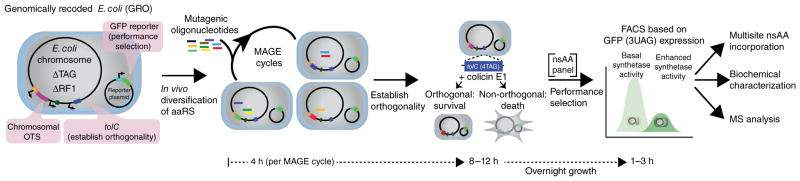
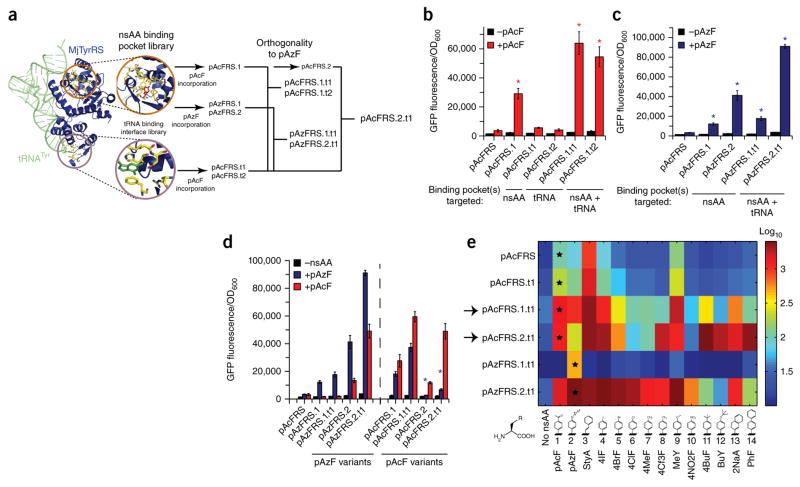
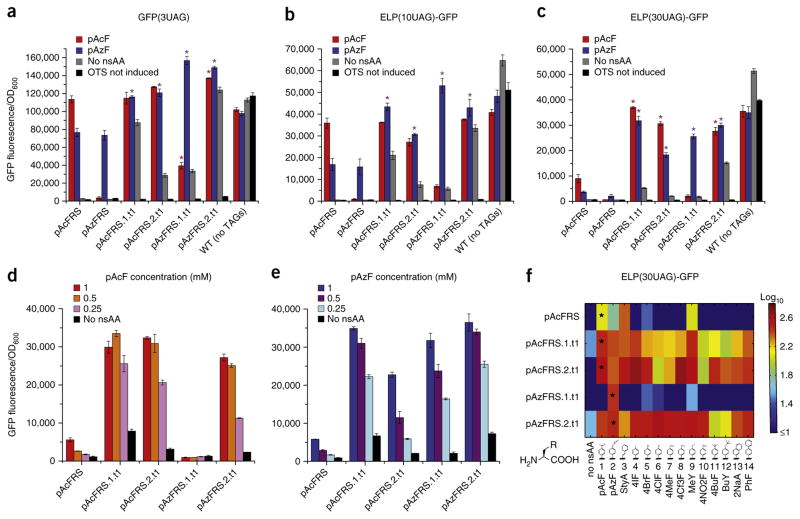
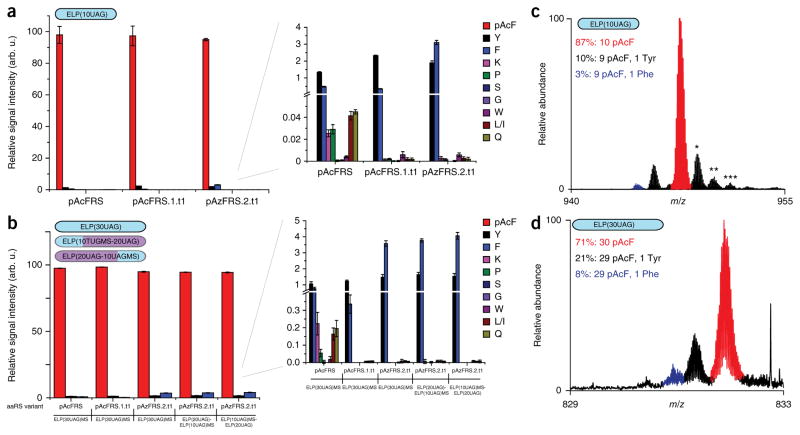
Expansion of the genetic code with nonstandard amino acids (nsAAs) has enabled biosynthesis of proteins with diverse new chemistries. However, this technology has been largely restricted to proteins containing a single or few nsAA instances. Here we describe an in vivo evolution approach in a genomically recoded Escherichia coli strain for the selection of orthogonal translation systems capable of multi-site nsAA incorporation. We evolved chromosomal aminoacyl-tRNA synthetases (aaRSs) with up to 25-fold increased protein production for p-acetyl-L-phenylalanine and p-azido-L-phenylalanine (pAzF). We also evolved aaRSs with tunable specificities for 14 nsAAs, including an enzyme that efficiently charges pAzF while excluding 237 other nsAAs. These variants enabled production of elastin-like-polypeptides with 30 nsAA residues at high yields (∼50 mg/L) and high accuracy of incorporation (>95%). This approach to aaRS evolution should accelerate and expand our ability to produce functionalized proteins and sequence-defined polymers with diverse chemistries.
Conflict of interest statement
The authors declare competing financial interests: details are available in the online version of the paper.
Figures
References
-
- Chin JW. Expanding and reprogramming the genetic code of cells and animals. Annu Rev Biochem. 2014;83:379–408. - PubMed
-
- Link AJ, Mock ML, Tirrell DA. Non-canonical amino acids in protein engineering. Curr Opin Biotechnol. 2003;14:603–609. - PubMed
-
- Liu CC, Schultz PG. Adding new chemistries to the genetic code. Annu Rev Biochem. 2010;79:413–444. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
