

Review
doi: 10.1039/c5np00112a.
Recent advances in the biosynthesis of unusual polyketide synthase substrates
Affiliations
- PMID: 26571143
- PMCID: PMC4742410
- DOI: 10.1039/c5np00112a
Item in Clipboard
Review
Recent advances in the biosynthesis of unusual polyketide synthase substrates
Nat Prod Rep.
2016 Feb.
Abstract
This highlight provides an overview of recent advances in understanding the diversity of polyketide synthase (PKS) substrate building blocks. Substrates functioning as starter units and extender units contribute significantly to the chemical complexity and structural diversity exhibited by this class of natural products. This article complements and extends upon the current comprehensive reviews that have been published on these two topics (Moore and Hertweck, Nat. Prod. Rep., 2002, 19, 70; Chan et al., Nat. Prod. Rep., 2009, 1, 90; Wilson and Moore, Nat. Prod. Rep., 2012, 29, 72).
Figures
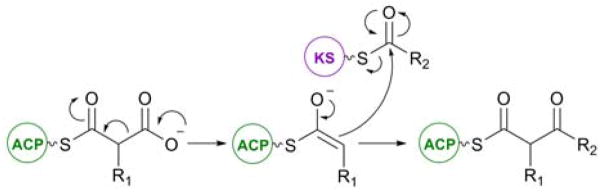
Mechanism of the decarboxylative Claisen condensation catalyzed by a KS domain during polyketide chain extension.
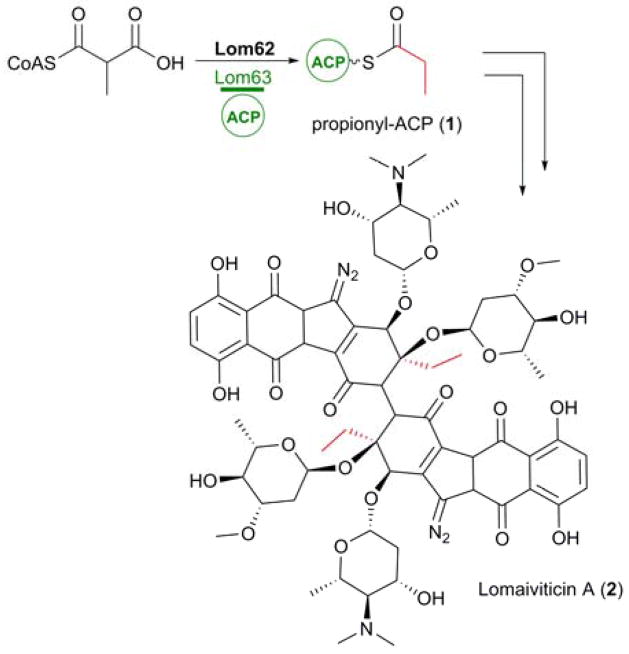
Biosynthesis of propionyl-ACP (1) via an unprecedented mechanism involving Lom62, a bifunctional AT/DC, for incorporation as a starter unit into lomaiviticin A (2).
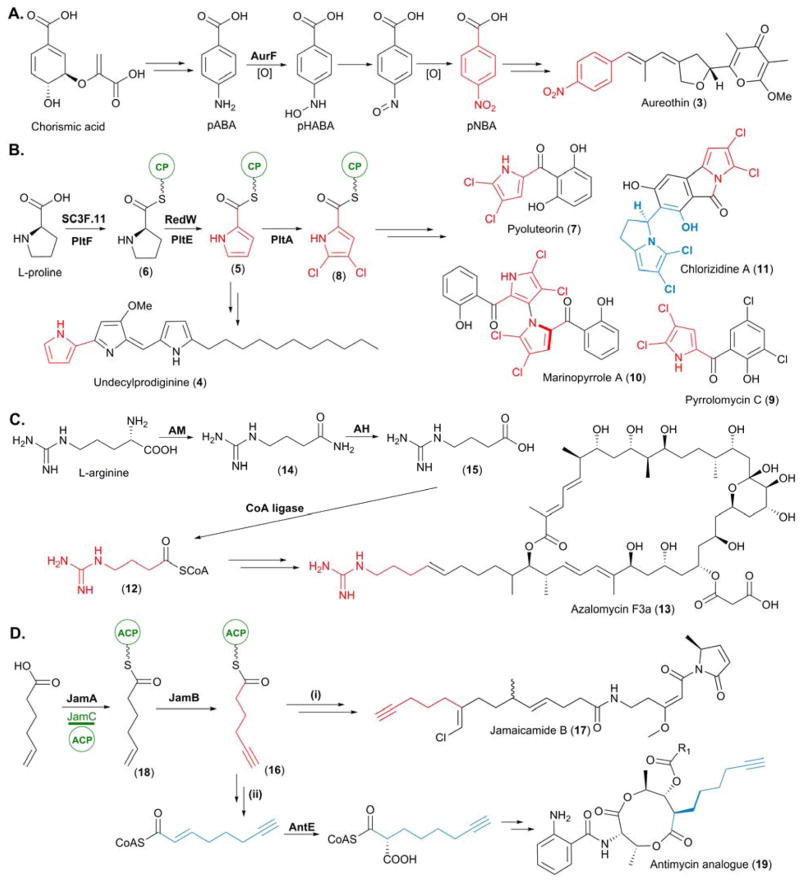
Biosynthesis of newly identified PKS starter units. (A) Biosynthesis of p-nitrobenzoic acid (pNBA), the starter unit for aureothin (3) biosynthesis. (B) Biosynthesis of the L-proline derived starter units pyrrolyl-CP (5) and 4,5-dichloropyrrolyl-CP (8). (C) Biosynthesis of the L-arginine derived starter unit 4-guanidinobutyryl-CoA (12). (D) Biosynthesis of 5-hexynoyl-ACP (16) for (I) incorporation into jamaicamide (17) and (II) utilization of its biosynthetic pathway for the bioengineering of an antimycin analogue (19). In this figure and all others in this article, we highlight the unusual starter unit and extender unit derived atoms in red and blue, respectively.
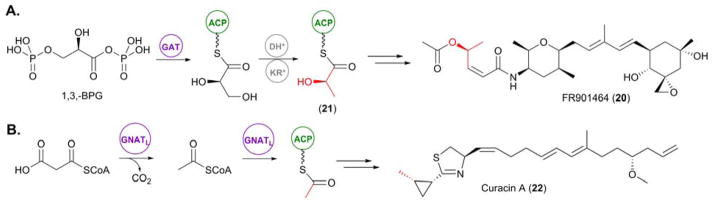
mechanisms for the incorporation of PKS starter units. (A) Incorporation of a glycyl moiety into FR901464 (20) via the unusual GAT domain located within the loading module for this PKS assembly line. (B) Priming of the curacin A (22) PKS via an unusual GNATL domain located within the initiating module of the PKS.
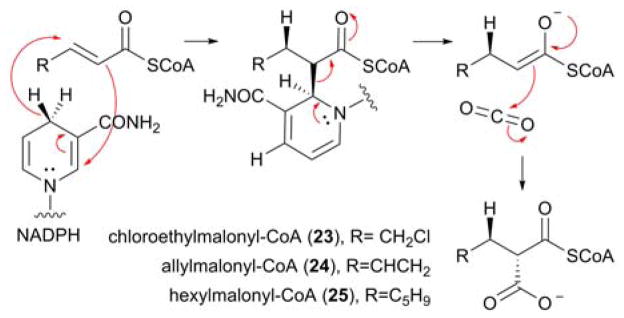
Proposed catalytic mechanism of the reductive carboxylation catalyzed by CCRs, with the regiosepecificity and the stereoselectivity of the reaction indicated. The structures of previously characterized atypical PKS extender units, biosynthesised by CCRs, are also highlighted.
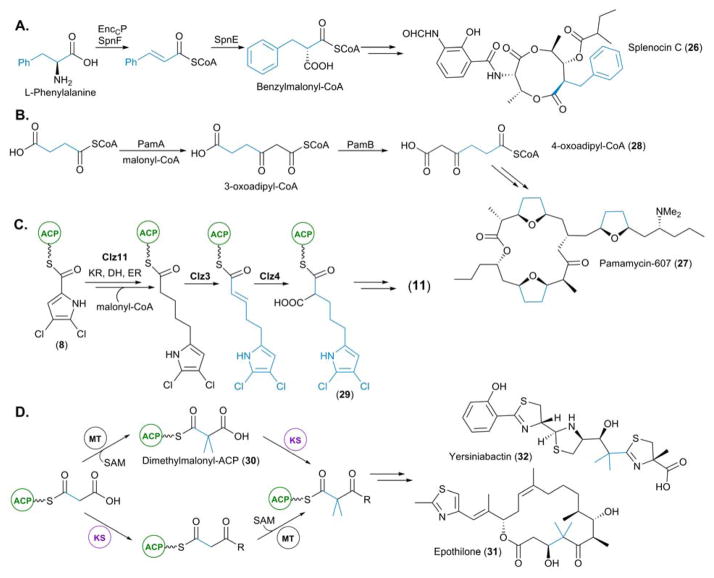
Biosynthesis of newly identified PKS extender units. (A) Biosynthesis of benzylmalonyl-CoA incorporated into the splenocins. (B) Biosynthesis of 3-oxoadipyl-CoA for incorporation into the pamamycins. (C) Biosynthesis of dichloropyrrolepropyl-ACP for incorporation into chlorizidine A (11, see blue highlight in Figure 3B). (D) Two characterized routes for the incorporation of gem-dimethyl groups present in yersinibactin and epothilone. The route exclusively utilized by EpoM8 (the epothilone PKS) proceeds via dimethylmalonyl-ACP (top pathway).
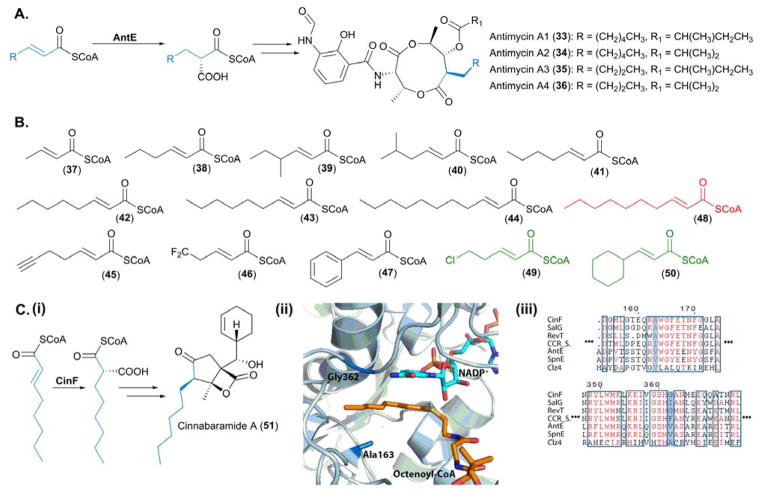
Biochemical and structural characterization of CCRs involved in PKS extender unit biosynthesis. (A) Native reactions catalyzed by AntE, the pathway specific CCR, involved in antimycin biosynthesis. (B) Substrates used to probe the promiscuity of AntE – the substrates in black were tolerated in vitro by AntE, substrates in green were tolerated in vivo by AntE and the malonate derivative highlighted in red was not accepted as a substrate. (Ci) Native reaction catalyzed by CinF, for incorporation of hexanoylmalonyl-CoA into cinnabaramide A. (Cii) Active site of CinF, crystallized in complex with both NADH and 2-octenoyl-CoA; residues essential for substrate specificity of CinF are highlighted. The PDB accession number for CinF with ligands bound is 4A0S and the image was generated in Pymol. (Ciii) Sequence alignments of characterized CCRs, with the residues conferring substrate specificity highlighted in blue. SalG is responsible for the biosynthesis of chloroethylmalonyl-CoA (23) in salinosporamide A biosynthesis, RevT is responsible for the biosynthesis of alkylmalonyl-CoAs incorporated into the reveromycins, CCR is from Streptomyces collinus and catalyzes the reductive decarboxylation of crotonyl-CoA into methylmalonyl-CoA, AntE is the highly promiscuous CCR from the antimycin pathway, SpnE is involved in the biosynthesis of benzylmalonyl-CoA from the splenocin biosynthetic gene cluster, and Clz4 is responsible for the biosynthesis of ACP-linked dichloropyrrolepropyl extender unit incorporated into chlorizidine A.
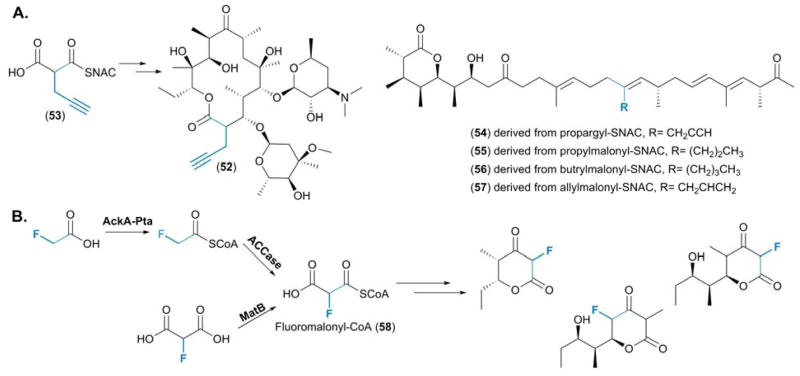
(A) Structures of the engineered polyketide 2-propargylerythromycin, resulting from the selection of 2-propargylmalonate by AT5DEBS, and the pre-monensin series, derived from the feeding a variety of “non-native” precursors for generation of novel derivatives. (B) The two distinct routes established for the provision of fluoromalonyl-CoA (58) and the suite of fluorinated polyketide molecules generated from the minimal DEBS system.
References
-
- Staunton J, Weissman KJ. Nat Prod Rep. 2001;18:380–416. - PubMed
-
- Fischbach MA, Walsh CT. Chem Rev. 2006;106:3468–3496. - PubMed
-
- Hertweck C, Luzhetskyy A, Rebets Y, Bechthold A. Nat Prod Rep. 2007;24:162–190. - PubMed
-
- Austin MB, Noel JP. Nat Prod Rep. 2003;20:79–110. - PubMed
-
- Hertweck C. Angew Chem, Int Ed. 2009;48:4688–4716. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous