

Branched-chain amino acid catabolism fuels adipocyte differentiation and lipogenesis
- PMID: 26571352
- PMCID: PMC4684771
- DOI: 10.1038/nchembio.1961
Branched-chain amino acid catabolism fuels adipocyte differentiation and lipogenesis
Abstract
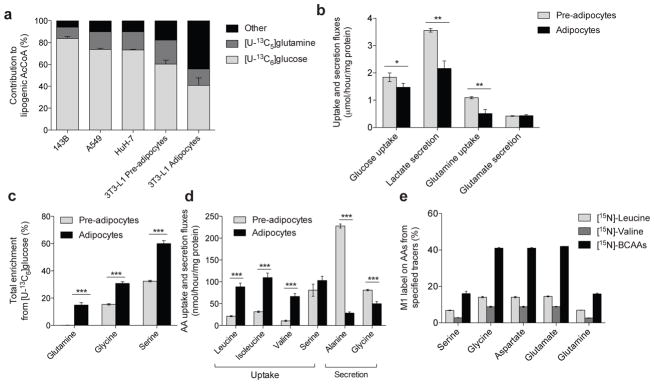
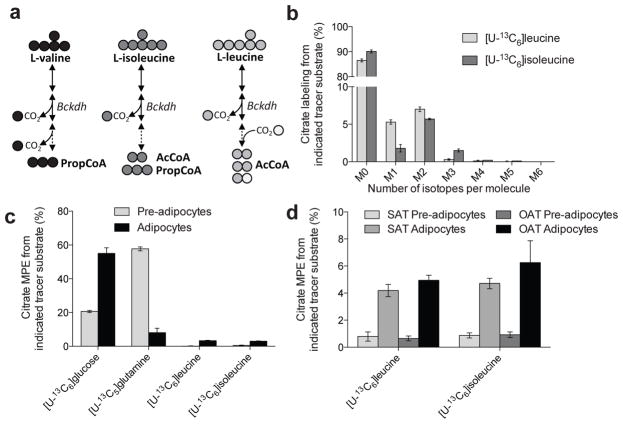
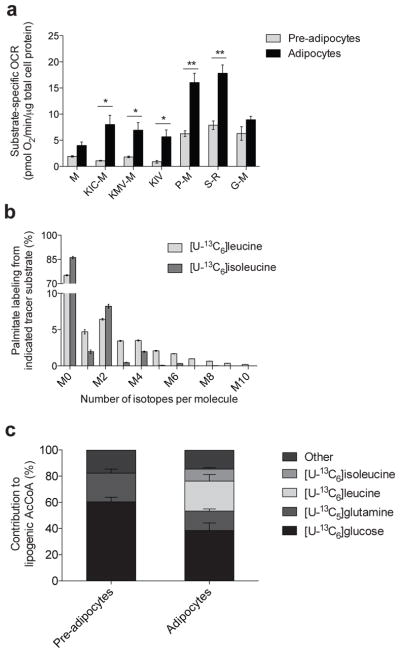
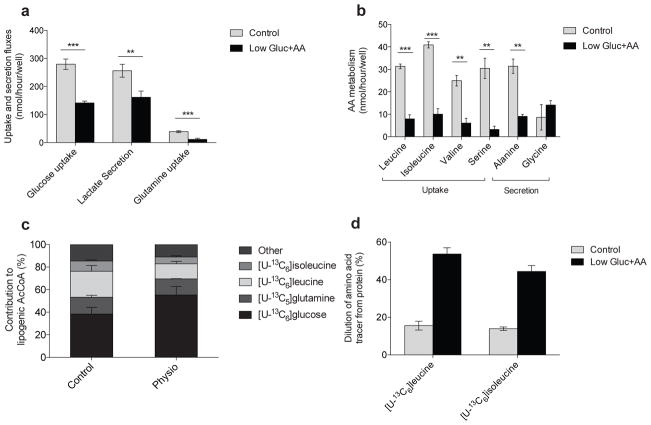
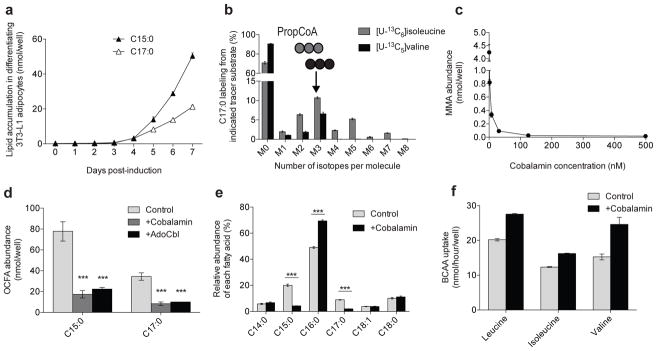
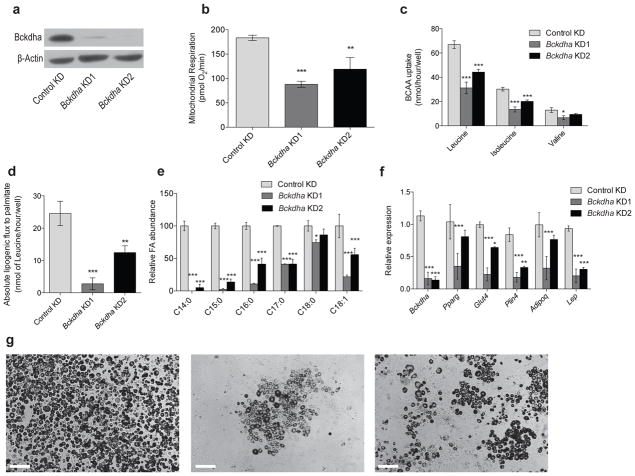
Adipose tissue plays important roles in regulating carbohydrate and lipid homeostasis, but less is known about the regulation of amino acid metabolism in adipocytes. Here we applied isotope tracing to pre-adipocytes and differentiated adipocytes to quantify the contributions of different substrates to tricarboxylic acid (TCA) metabolism and lipogenesis. In contrast to proliferating cells, which use glucose and glutamine for acetyl-coenzyme A (AcCoA) generation, differentiated adipocytes showed increased branched-chain amino acid (BCAA) catabolic flux such that leucine and isoleucine from medium and/or from protein catabolism accounted for as much as 30% of lipogenic AcCoA pools. Medium cobalamin deficiency caused methylmalonic acid accumulation and odd-chain fatty acid synthesis. Vitamin B12 supplementation reduced these metabolites and altered the balance of substrates entering mitochondria. Finally, inhibition of BCAA catabolism compromised adipogenesis. These results quantitatively highlight the contribution of BCAAs to adipocyte metabolism and suggest that BCAA catabolism has a functional role in adipocyte differentiation.
Conflict of interest statement
No potential conflicts of interest relevant to this article were reported.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
