

Pericentromeric satellite repeat expansions through RNA-derived DNA intermediates in cancer
- PMID: 26575630
- PMCID: PMC4679016
- DOI: 10.1073/pnas.1518008112
Pericentromeric satellite repeat expansions through RNA-derived DNA intermediates in cancer
Abstract
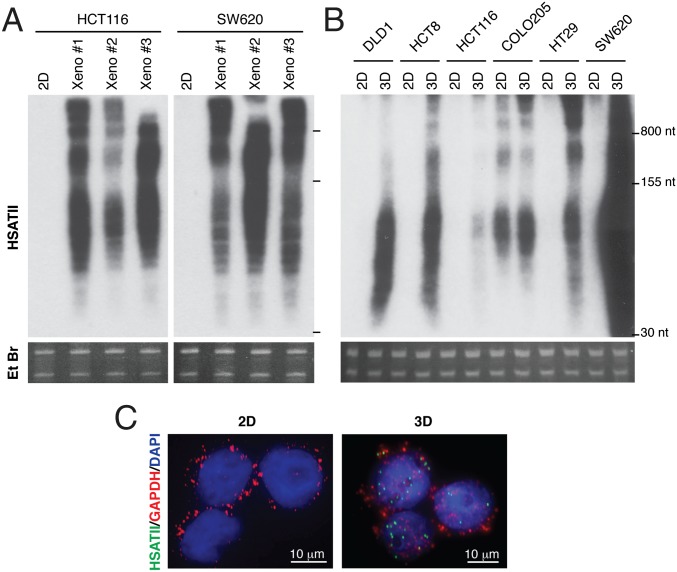
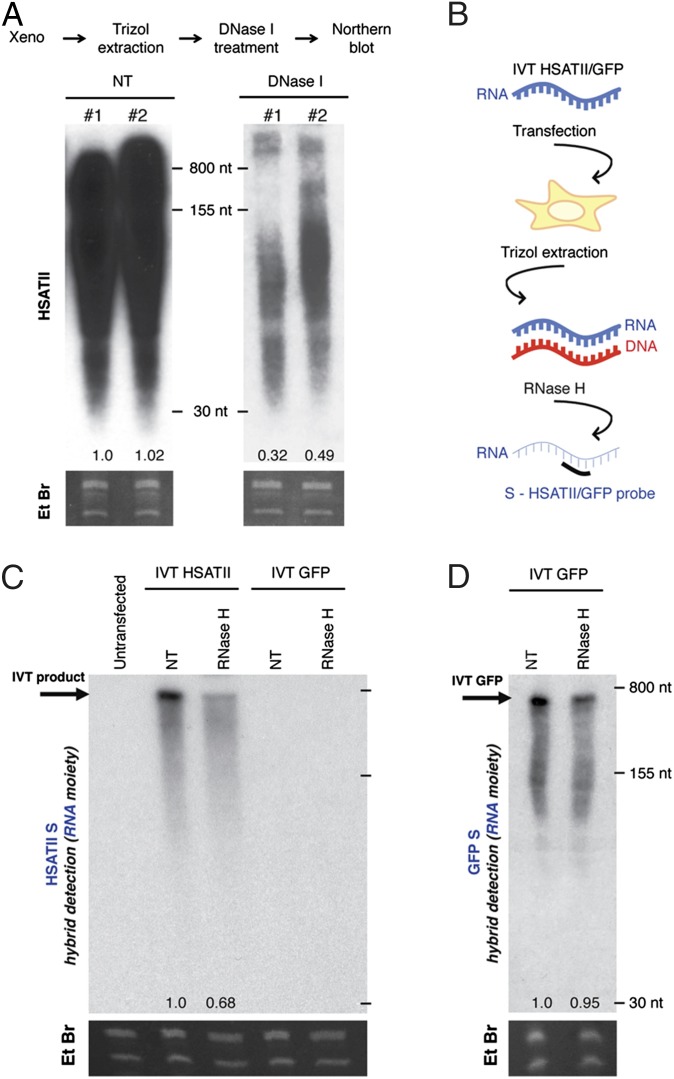
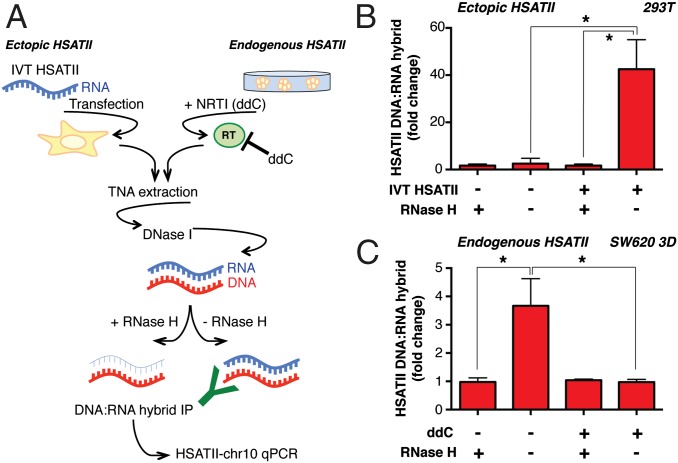
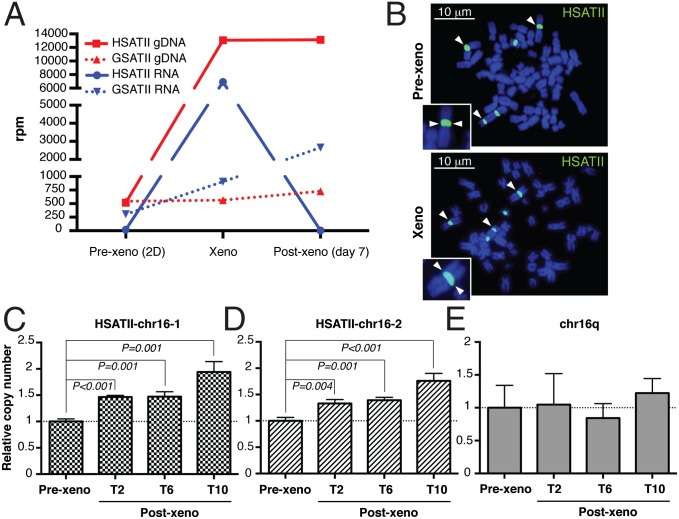
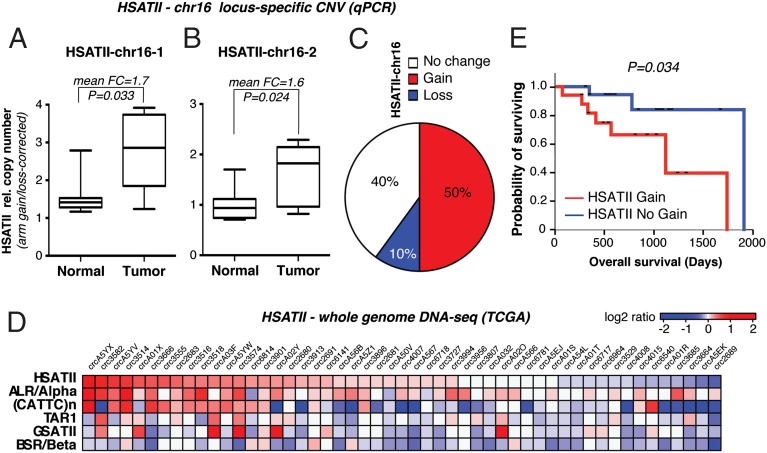
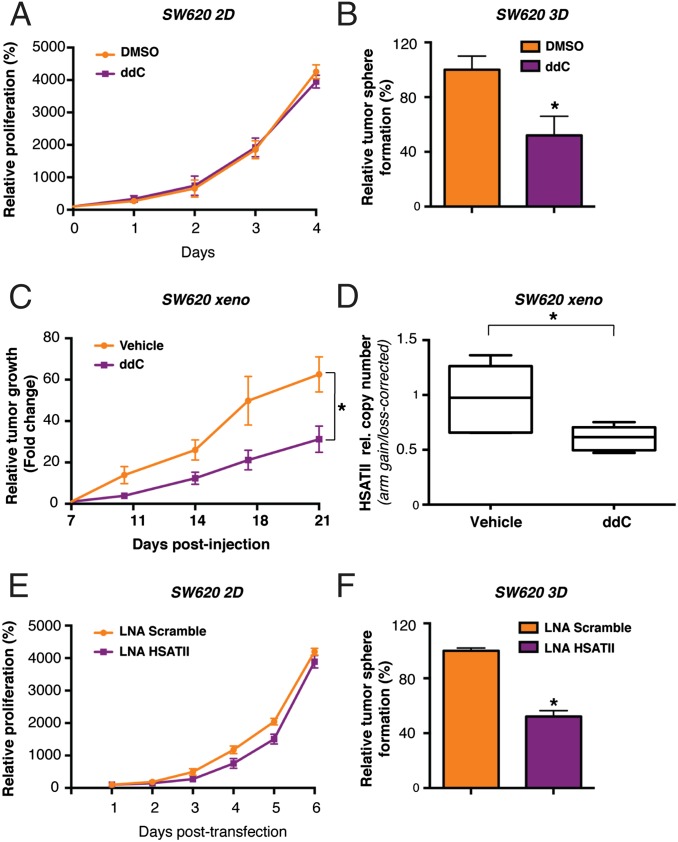
Aberrant transcription of the pericentromeric human satellite II (HSATII) repeat is present in a wide variety of epithelial cancers. In deriving experimental systems to study its deregulation, we observed that HSATII expression is induced in colon cancer cells cultured as xenografts or under nonadherent conditions in vitro, but it is rapidly lost in standard 2D cultures. Unexpectedly, physiological induction of endogenous HSATII RNA, as well as introduction of synthetic HSATII transcripts, generated cDNA intermediates in the form of DNA/RNA hybrids. Single molecule sequencing of tumor xenografts showed that HSATII RNA-derived DNA (rdDNA) molecules are stably incorporated within pericentromeric loci. Suppression of RT activity using small molecule inhibitors reduced HSATII copy gain. Analysis of whole-genome sequencing data revealed that HSATII copy number gain is a common feature in primary human colon tumors and is associated with a lower overall survival. Together, our observations suggest that cancer-associated derepression of specific repetitive sequences can promote their RNA-driven genomic expansion, with potential implications on pericentromeric architecture.
Keywords: cancer; repeats; reverse transcription; satellites.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Silent pericentromeric repeats speak out.Proc Natl Acad Sci U S A. 2015 Dec 8;112(49):15008-9. doi: 10.1073/pnas.1520341112. Epub 2015 Nov 18. Proc Natl Acad Sci U S A. 2015. PMID: 26582791 Free PMC article. No abstract available.
References
-
- Eymery A, Callanan M, Vourc’h C. The secret message of heterochromatin: New insights into the mechanisms and function of centromeric and pericentric repeat sequence transcription. Int J Dev Biol. 2009;53(2-3):259–268. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
